

Mutagenesis of the Peptide Inhibitor of ASIC3 Channel Introduces Binding to Thumb Domain of ASIC1a but Reduces Analgesic Activity
- PMID: 39330263
- PMCID: PMC11432795
- DOI: 10.3390/md22090382
Mutagenesis of the Peptide Inhibitor of ASIC3 Channel Introduces Binding to Thumb Domain of ASIC1a but Reduces Analgesic Activity
Abstract
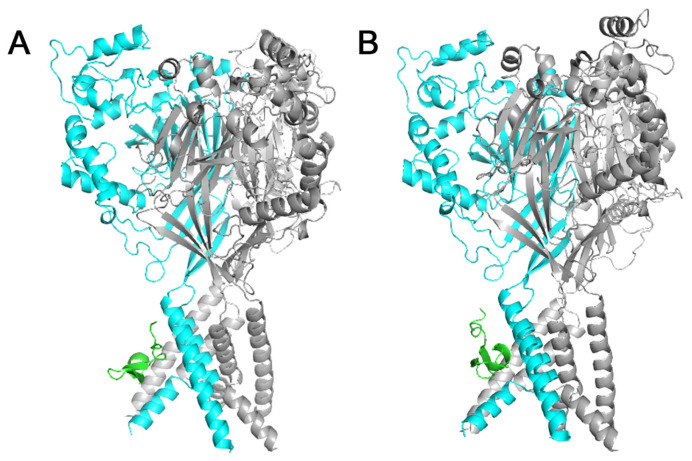
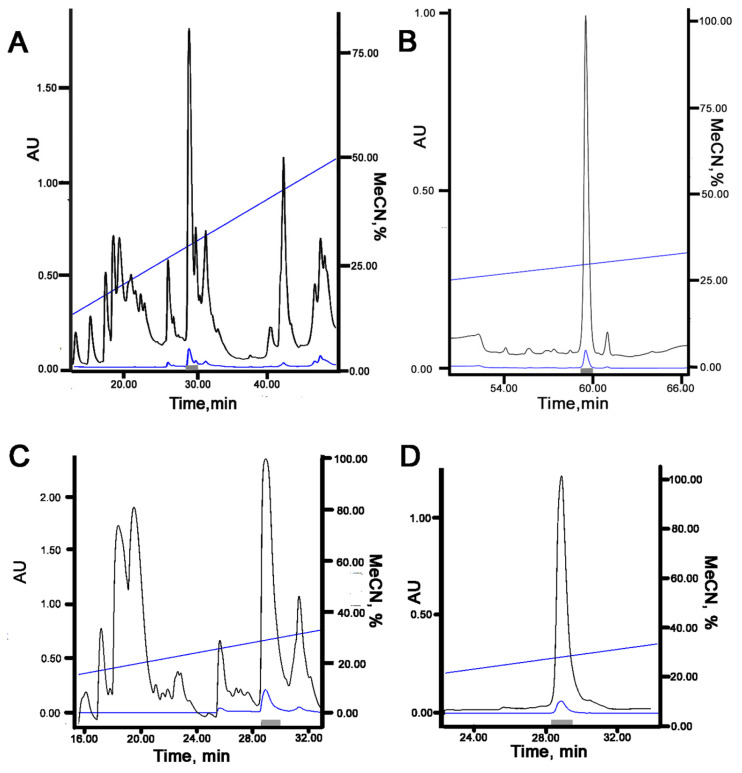
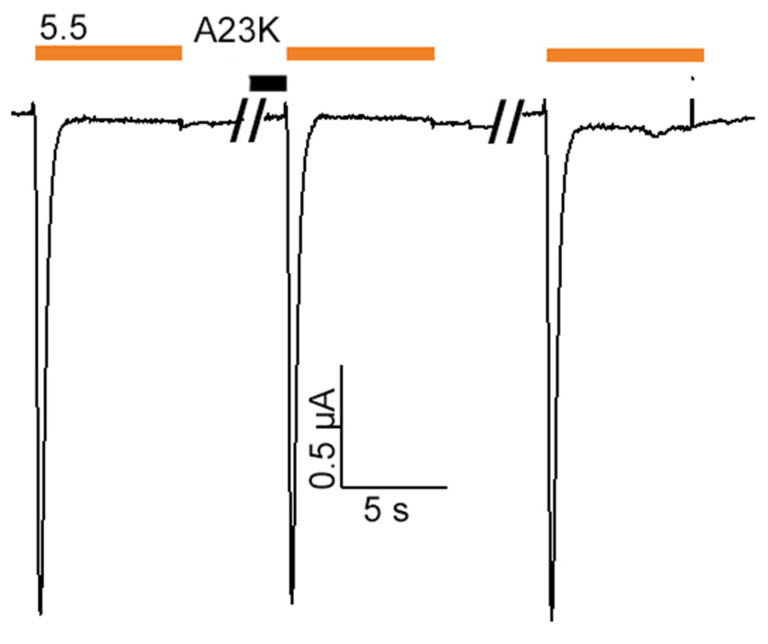
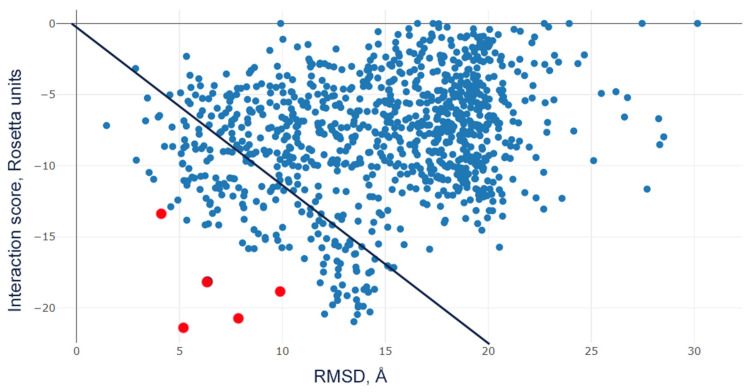
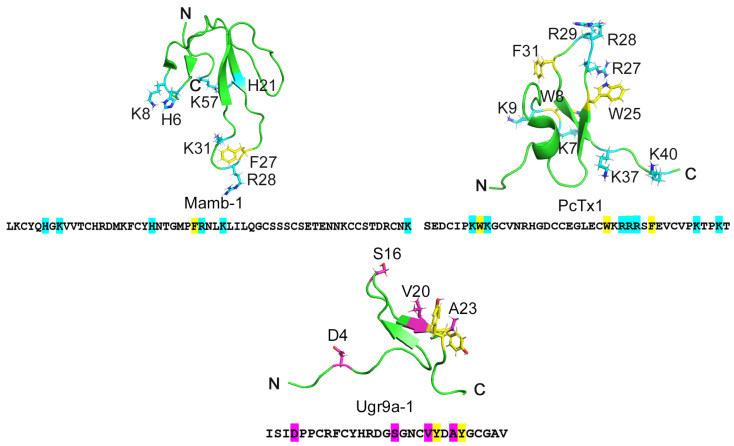
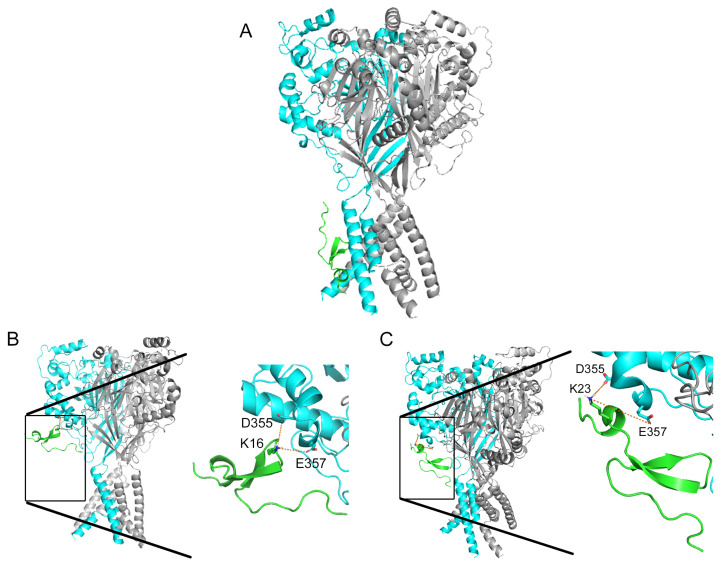
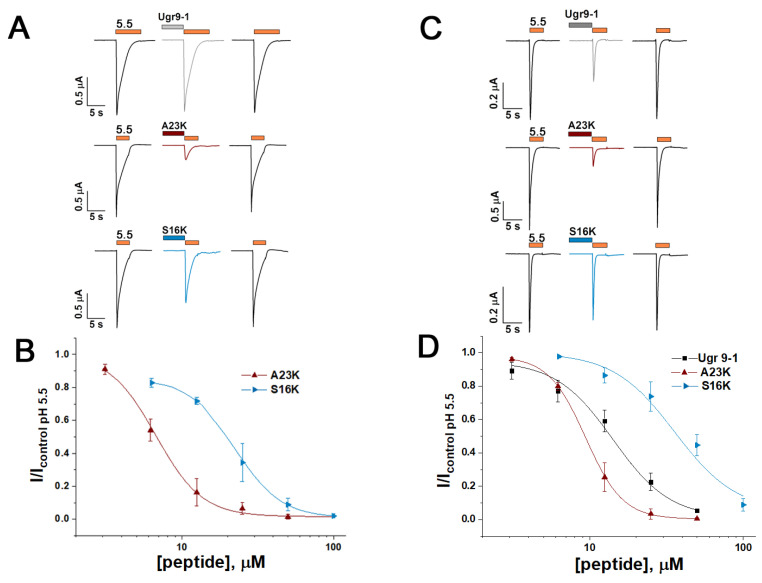
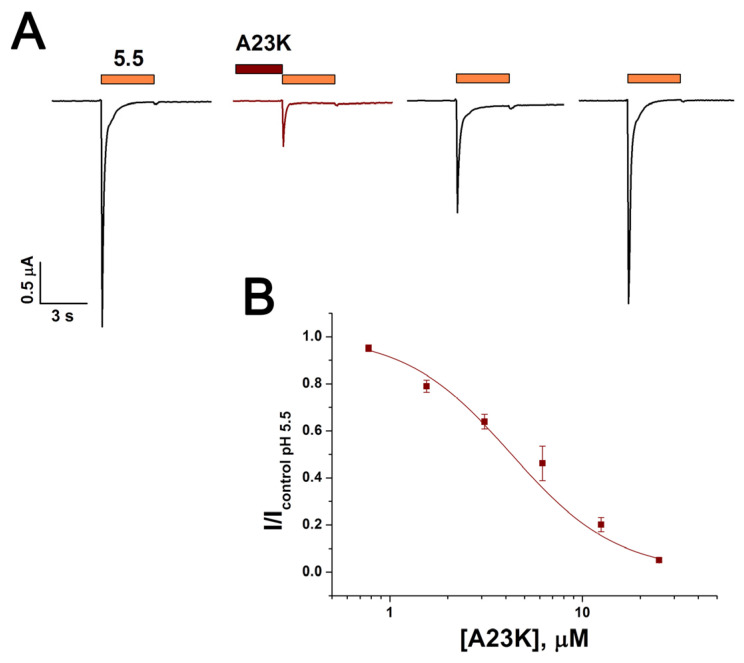
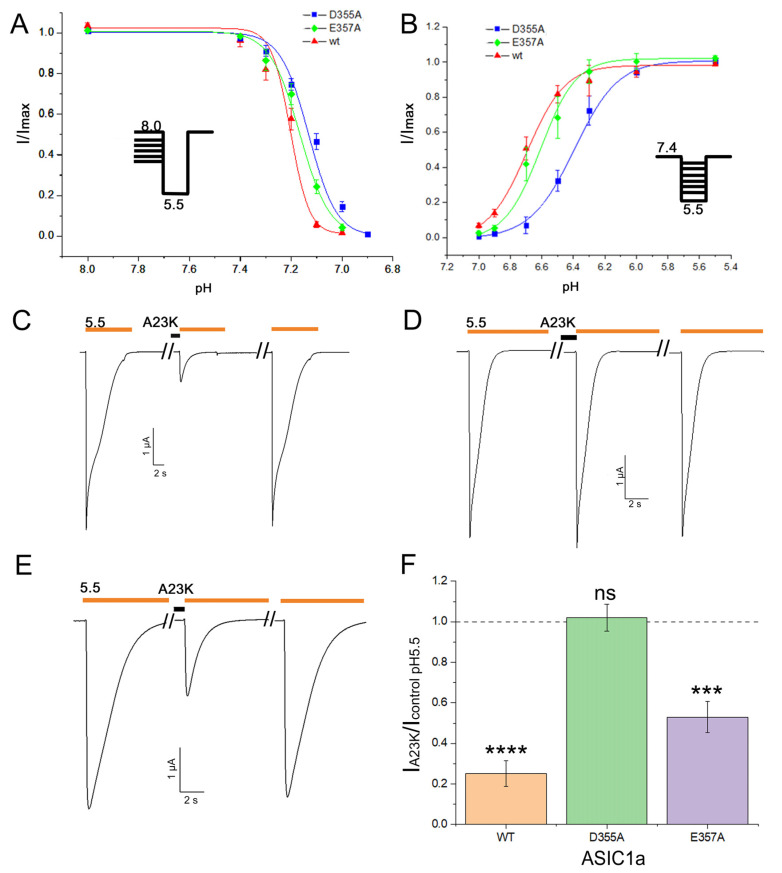
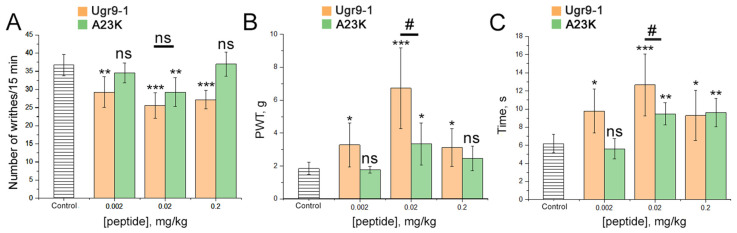
Acid-sensing ion channels (ASICs), which act as proton-gating sodium channels, have garnered attention as pharmacological targets. ASIC1a isoform, notably prevalent in the central nervous system, plays an important role in synaptic plasticity, anxiety, neurodegeneration, etc. In the peripheral nervous system, ASIC1a shares prominence with ASIC3, the latter well established for its involvement in pain signaling, mechanical sensitivity, and inflammatory hyperalgesia. However, the precise contributions of ASIC1a in peripheral functions necessitate thorough investigation. To dissect the specific roles of ASICs, peptide ligands capable of modulating these channels serve as indispensable tools. Employing molecular modeling, we designed the peptide targeting ASIC1a channel from the sea anemone peptide Ugr9-1, originally targeting ASIC3. This peptide (A23K) retained an inhibitory effect on ASIC3 (IC50 9.39 µM) and exhibited an additional inhibitory effect on ASIC1a (IC50 6.72 µM) in electrophysiological experiments. A crucial interaction between the Lys23 residue of the A23K peptide and the Asp355 residue in the thumb domain of the ASIC1a channel predicted by molecular modeling was confirmed by site-directed mutagenesis of the channel. However, A23K peptide revealed a significant decrease in or loss of analgesic properties when compared to the wild-type Ugr9-1. In summary, using A23K, we show that negative modulation of the ASIC1a channel in the peripheral nervous system can compromise the efficacy of an analgesic drug. These results provide a compelling illustration of the complex balance required when developing peripheral pain treatments targeting ASICs.
Keywords: acid-sensing ion channel; analgesic effect; molecular docking; mutagenesis; pain models; peptide ligand.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Analgesic Activity of Acid-Sensing Ion Channel 3 (ASIС3) Inhibitors: Sea Anemones Peptides Ugr9-1 and APETx2 versus Low Molecular Weight Compounds.Mar Drugs. 2018 Dec 12;16(12):500. doi: 10.3390/md16120500. Mar Drugs. 2018. PMID: 30545037 Free PMC article.
-
Spider-derived peptide LCTx-F2 suppresses ASIC channels by occupying the acidic pocket.J Biol Chem. 2025 Mar;301(3):108286. doi: 10.1016/j.jbc.2025.108286. Epub 2025 Feb 10. J Biol Chem. 2025. PMID: 39938802 Free PMC article.
-
Peptides inhibitors of acid-sensing ion channels.Toxicon. 2007 Feb;49(2):271-84. doi: 10.1016/j.toxicon.2006.09.026. Epub 2006 Oct 4. Toxicon. 2007. PMID: 17113616 Review.
-
Pharmacological modulation of Acid-Sensing Ion Channels 1a and 3 by amiloride and 2-guanidine-4-methylquinazoline (GMQ).Neuropharmacology. 2017 Oct;125:429-440. doi: 10.1016/j.neuropharm.2017.08.004. Epub 2017 Aug 9. Neuropharmacology. 2017. PMID: 28802647
-
Research strategies for pain in lumbar radiculopathy focusing on acid-sensing ion channels and their toxins.Curr Top Med Chem. 2015;15(7):617-30. doi: 10.2174/1568026615666150217112652. Curr Top Med Chem. 2015. PMID: 25686734 Review.
Cited by
-
Two Amino Acid Substitutions Improve the Pharmacological Profile of the Snake Venom Peptide Mambalgin.Toxins (Basel). 2025 Feb 21;17(3):101. doi: 10.3390/toxins17030101. Toxins (Basel). 2025. PMID: 40137874 Free PMC article.
References
-
- Wemmie J.A., Chen J., Askwith C.C., Hruska-Hageman A.M., Price M.P., Nolan B.C., Yoder P.G., Lamani E., Hoshi T., Freeman J.H., et al. The Acid-Activated Ion Channel ASIC Contributes to Synaptic Plasticity, Learning, and Memory. Neuron. 2002;34:463–477. doi: 10.1016/S0896-6273(02)00661-X. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
