

Subcellular activation of β-adrenergic receptors using a spatially restricted antagonist
- PMID: 39331410
- PMCID: PMC11459184
- DOI: 10.1073/pnas.2404243121
Subcellular activation of β-adrenergic receptors using a spatially restricted antagonist
Abstract
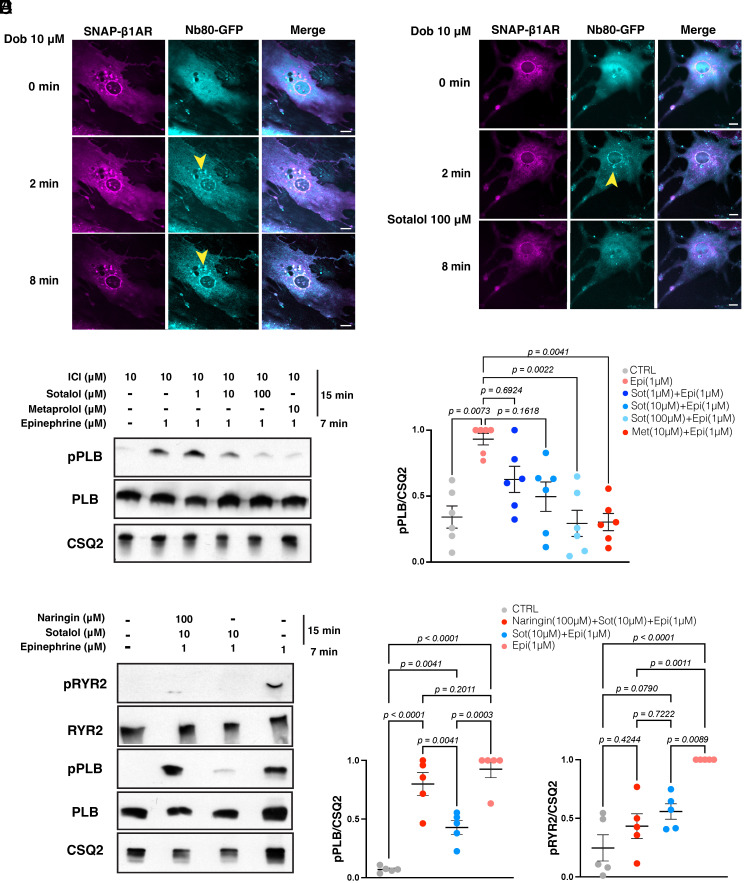
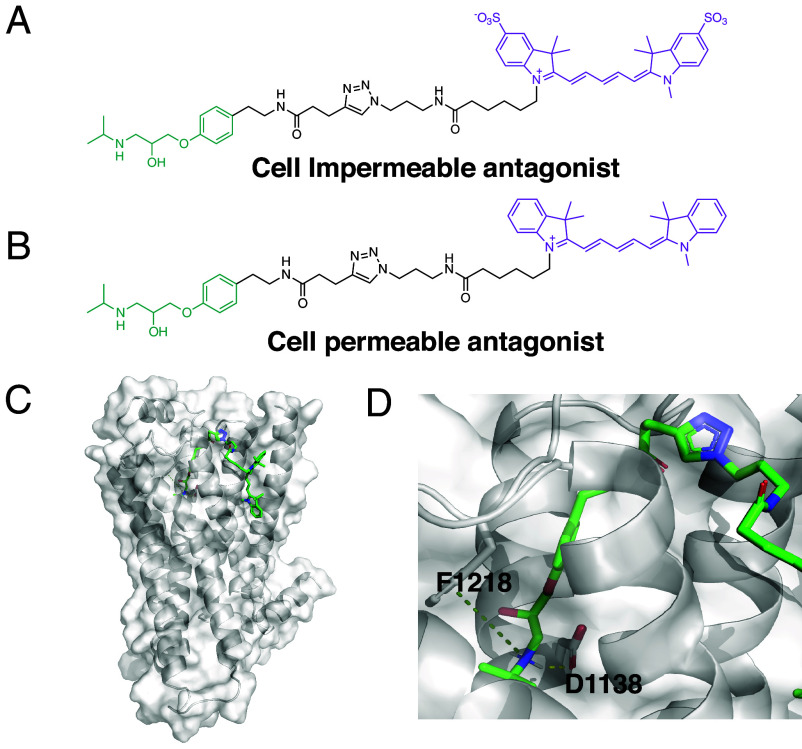
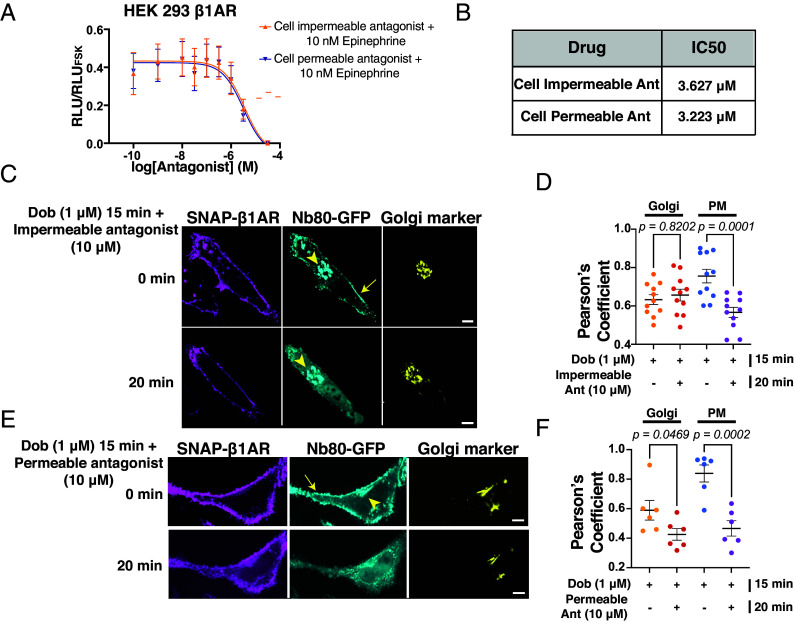
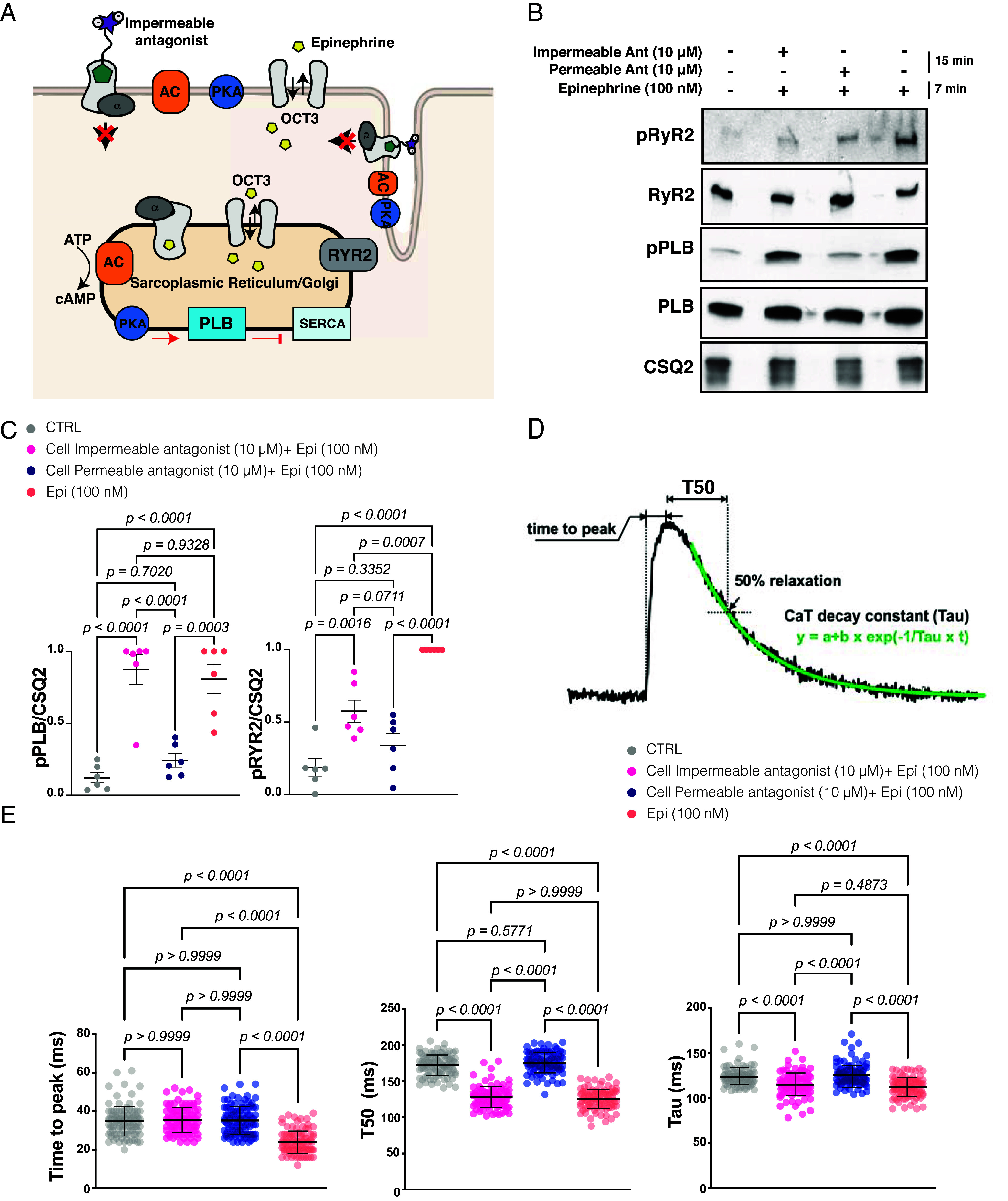
Gprotein-coupled receptors (GPCRs) regulate several physiological and pathological processes and represent the target of approximately 30% of Food and Drug Administration-approved drugs. GPCR-mediated signaling was thought to occur exclusively at the plasma membrane. However, recent studies have unveiled their presence and function at subcellular membrane compartments. There is a growing interest in studying compartmentalized signaling of GPCRs. This requires development of tools to separate GPCR signaling at the plasma membrane from the ones initiated at intracellular compartments. We leveraged the structural and pharmacological information available for β-adrenergic receptors (βARs) and focused on β1AR as exemplary GPCR that functions at subcellular compartments, and rationally designed spatially restricted antagonists. We generated a cell-impermeable βAR antagonist by conjugating a suitable pharmacophore to a sulfonate-containing fluorophore. This cell-impermeable antagonist only inhibited β1AR on the plasma membrane. In contrast, a cell-permeable βAR antagonist containing a nonsulfonated fluorophore efficiently inhibited both the plasma membrane and Golgi pools of β1ARs. Furthermore, the cell-impermeable antagonist selectively inhibited the phosphorylation of PKA downstream effectors near the plasma membrane, which regulate sarcoplasmic reticulum (SR) Ca2+ release in adult cardiomyocytes, while the β1AR Golgi pool remained active. Our tools offer promising avenues for investigating compartmentalized βAR signaling in various contexts, potentially advancing our understanding of βAR-mediated cellular responses in health and disease. They also offer a general strategy to study compartmentalized signaling for other GPCRs in various biological systems.
Keywords: GPCR signaling; Pharmacology; drug design.
Conflict of interest statement
Competing interests statement:K.M.S. has consulting agreements for the following companies, which involve monetary and/or stock compensation: Revolution Medicines, Black Diamond Therapeutics, BridGene Biosciences, Denali Therapeutics, Dice Molecules, eFFECTOR Therapeutics, Erasca, Genentech/Roche, Janssen Pharmaceuticals, Kumquat Biosciences, Kura Oncology, Mitokinin, Nested, Type6 Therapeutics, Venthera, Wellspring Biosciences (Araxes Pharma), Turning Point, Ikena, Initial Therapeutics, Vevo and BioTheryX.
Figures
Update of
-
Selective activation of intracellular β1AR using a spatially restricted antagonist.bioRxiv [Preprint]. 2023 Nov 22:2023.11.22.568314. doi: 10.1101/2023.11.22.568314. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 Oct;121(40):e2404243121. doi: 10.1073/pnas.2404243121. PMID: 38045405 Free PMC article. Updated. Preprint.
References
-
- Pierce K. L., Premont R. T., Lefkowitz R. J., Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 3, 639–650 (2002). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
