

New insights into the ovulatory process in the human ovary
- PMID: 39331957
- PMCID: PMC11696709
- DOI: 10.1093/humupd/dmae027
New insights into the ovulatory process in the human ovary
Abstract
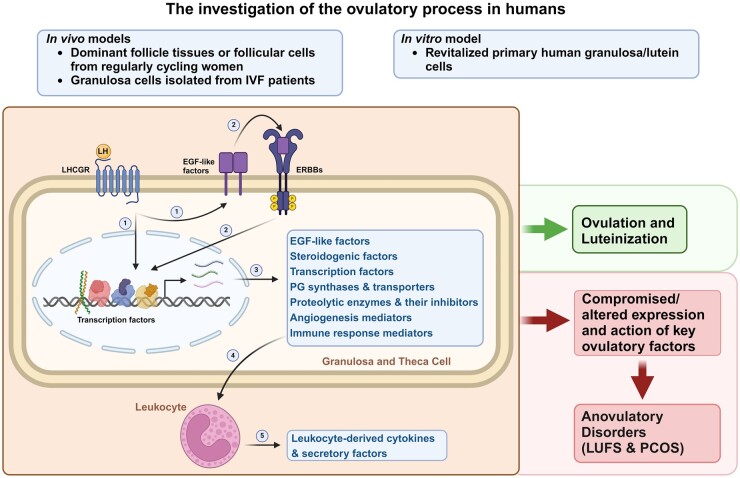
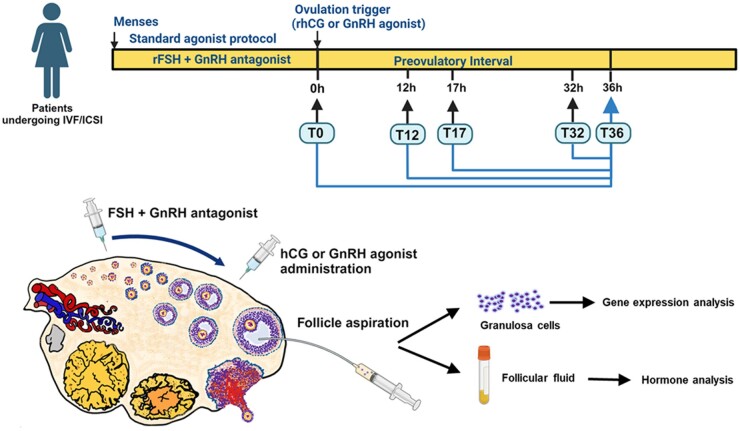
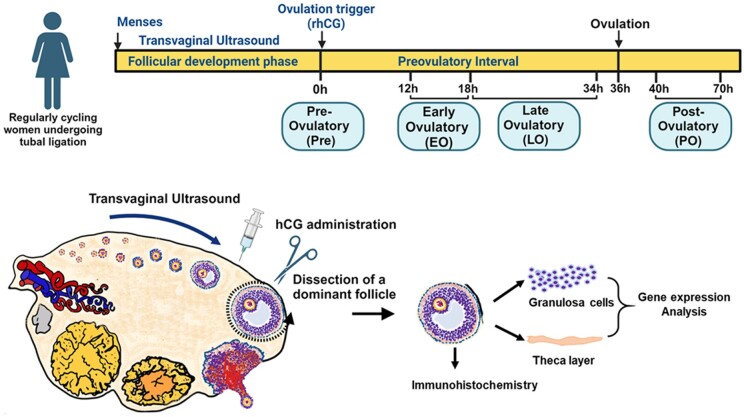
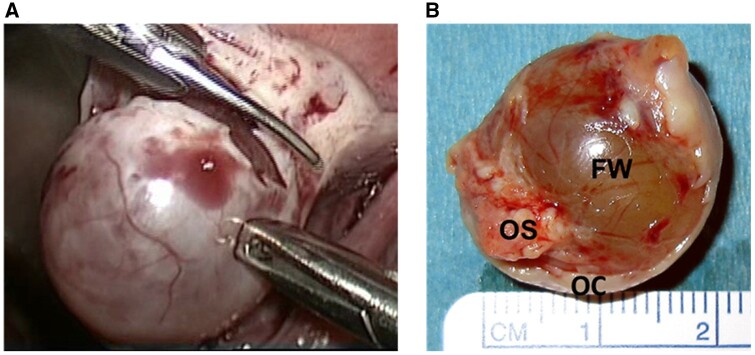
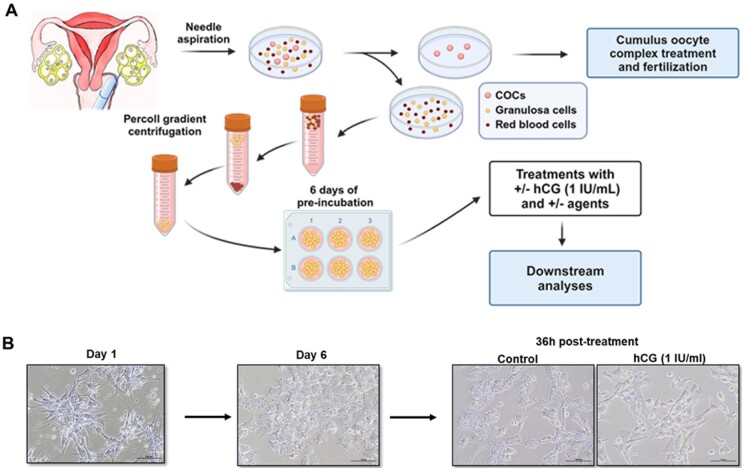
Background: Successful ovulation is essential for natural conception and fertility. Defects in the ovulatory process are associated with various conditions of infertility or subfertility in women. However, our understanding of the intra-ovarian biochemical mechanisms underlying this process in women has lagged compared to our understanding of animal models. This has been largely due to the limited availability of human ovarian samples that can be used to examine changes across the ovulatory period and delineate the underlying cellular/molecular mechanisms in women. Despite this challenge, steady progress has been made to improve our knowledge of the ovulatory process in women by: (i) collecting granulosa cells across the IVF interval, (ii) creating a novel approach to collecting follicular cells and tissues across the periovulatory period from normally cycling women, and (iii) developing unique in vitro models to examine the LH surge or hCG administration-induced ovulatory changes in gene expression, the regulatory mechanisms underlying the ovulatory changes, and the specific functions of the ovulatory factors.
Objective and rationale: The objective of this review is to summarize findings generated using in vivo and in vitro models of human ovulation, with the goal of providing new insights into the mechanisms underlying the ovulatory process in women.
Search methods: This review is based on the authors' own studies and a search of the relevant literature on human ovulation to date using PubMed search terms such as 'human ovulation EGF-signaling', 'human ovulation steroidogenesis', 'human ovulation transcription factor', 'human ovulation prostaglandin', 'human ovulation proteinase', 'human ovulation angiogenesis' 'human ovulation chemokine', 'human ovulatory disorder', 'human granulosa cell culture'. Our approach includes comparing the data from the authors' studies with the existing microarray or RNA-seq datasets generated using ovarian cells obtained throughout the ovulatory period from humans, monkeys, and mice.
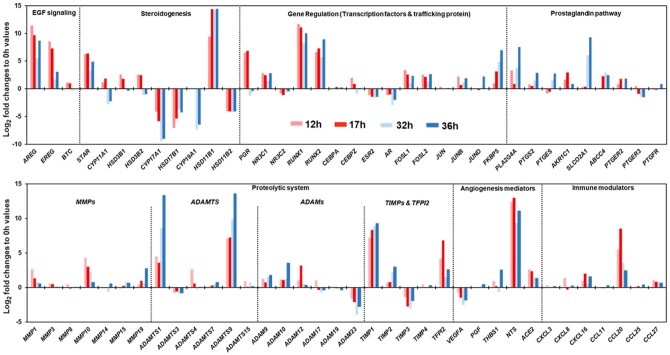
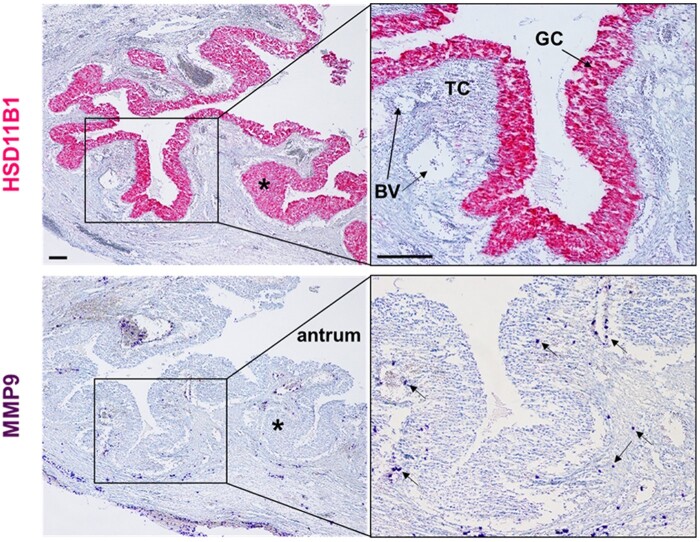
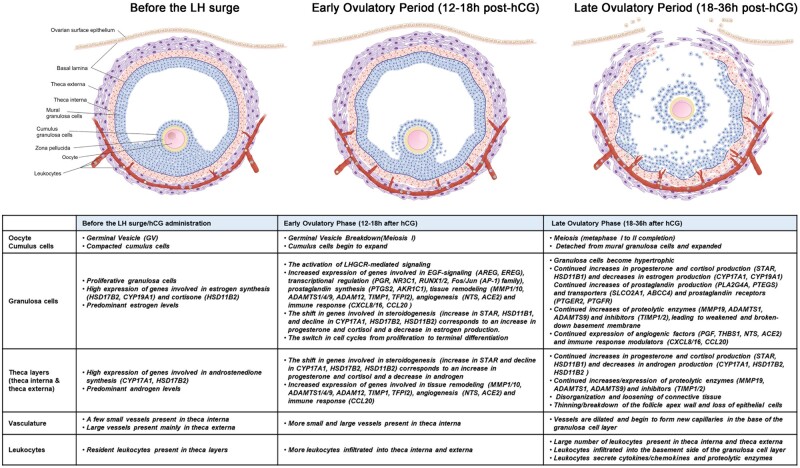
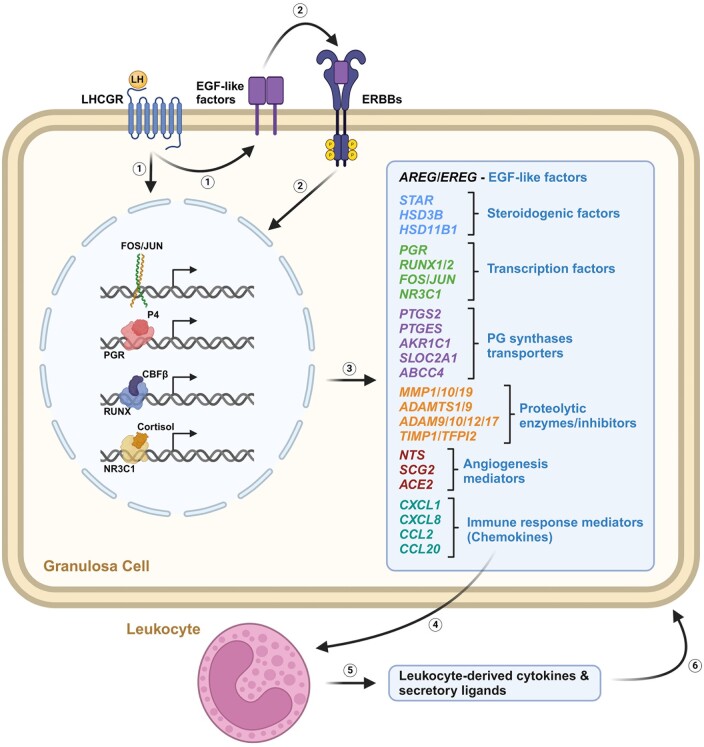
Outcomes: Current findings from studies using in vivo and in vitro models demonstrate that the LH surge or hCG administration increases the expression of ovulatory mediators, including EGF-like factors, steroids, transcription factors, prostaglandins, proteolytic systems, and other autocrine and paracrine factors, similar to those observed in other animal models such as rodents, ruminants, and monkeys. However, the specific ovulatory factors induced, their expression pattern, and their regulatory mechanisms vary among different species. These species-specific differences stress the necessity of utilizing human samples to delineate the mechanisms underlying the ovulatory process in women.
Wider implications: The data from human ovulation in vivo and in vitro models have begun to fill the gaps in our understanding of the ovulatory process in women. Further efforts are needed to discover novel ovulatory factors. One approach to address these gaps is to improve existing in vitro models to more closely mimic in vivo ovulatory conditions in humans. This is critically important as the knowledge obtained from these human studies can be translated directly to aid in the diagnosis of ovulation-associated pathological conditions, for the development of more effective treatment to help women with anovulatory infertility or, conversely, to better manage ovulation for contraceptive purposes.
Registration number: N/A.
Keywords: follicle; granulosa cells; infertility; oocyte; ovulation.
© The Author(s) 2024. Published by Oxford University Press on behalf of European Society of Human Reproduction and Embryology.
Conflict of interest statement
None declared.
Figures
References
-
- Akison LK, Robertson SA, Gonzalez MB, Richards JS, Smith CW, Russell DL, Robker RL. Regulation of the ovarian inflammatory response at ovulation by nuclear progesterone receptor. Am J Reprod Immunol 2018;79:e12835. - PubMed
-
- Andersen CY, Morineau G, Fukuda M, Westergaard LG, Ingerslev HJ, Fiet J, Byskov AG. Assessment of the follicular cortisol:cortisone ratio. Hum Reprod 1999;14:1563–1568. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
