

Evolution of novel sensory organs in fish with legs
- PMID: 39332400
- PMCID: PMC11552235
- DOI: 10.1016/j.cub.2024.08.014
Evolution of novel sensory organs in fish with legs
Abstract
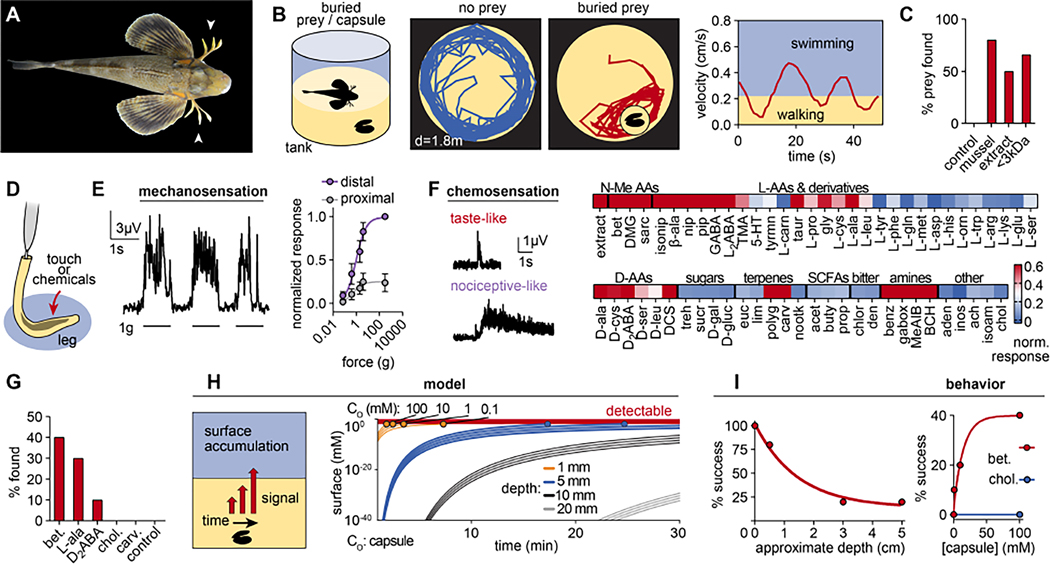
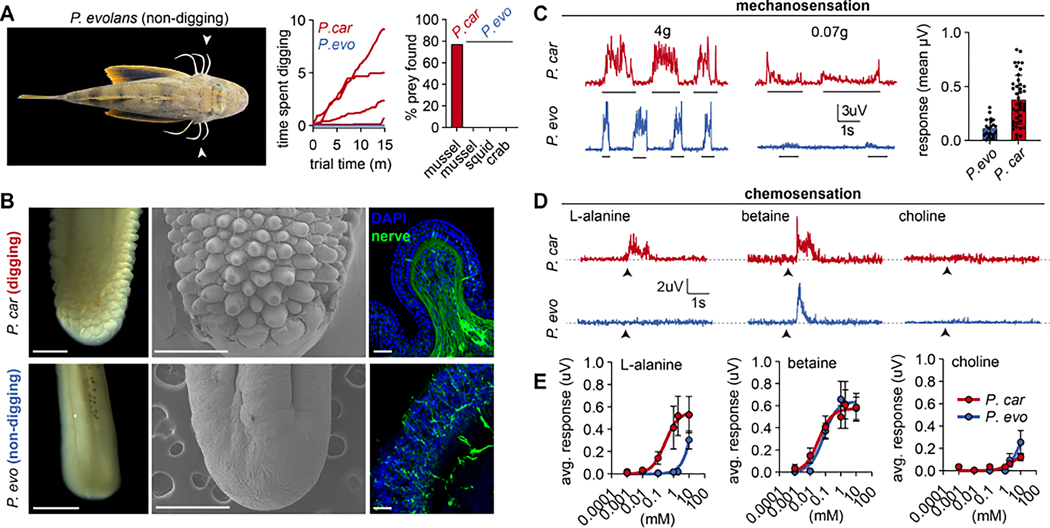
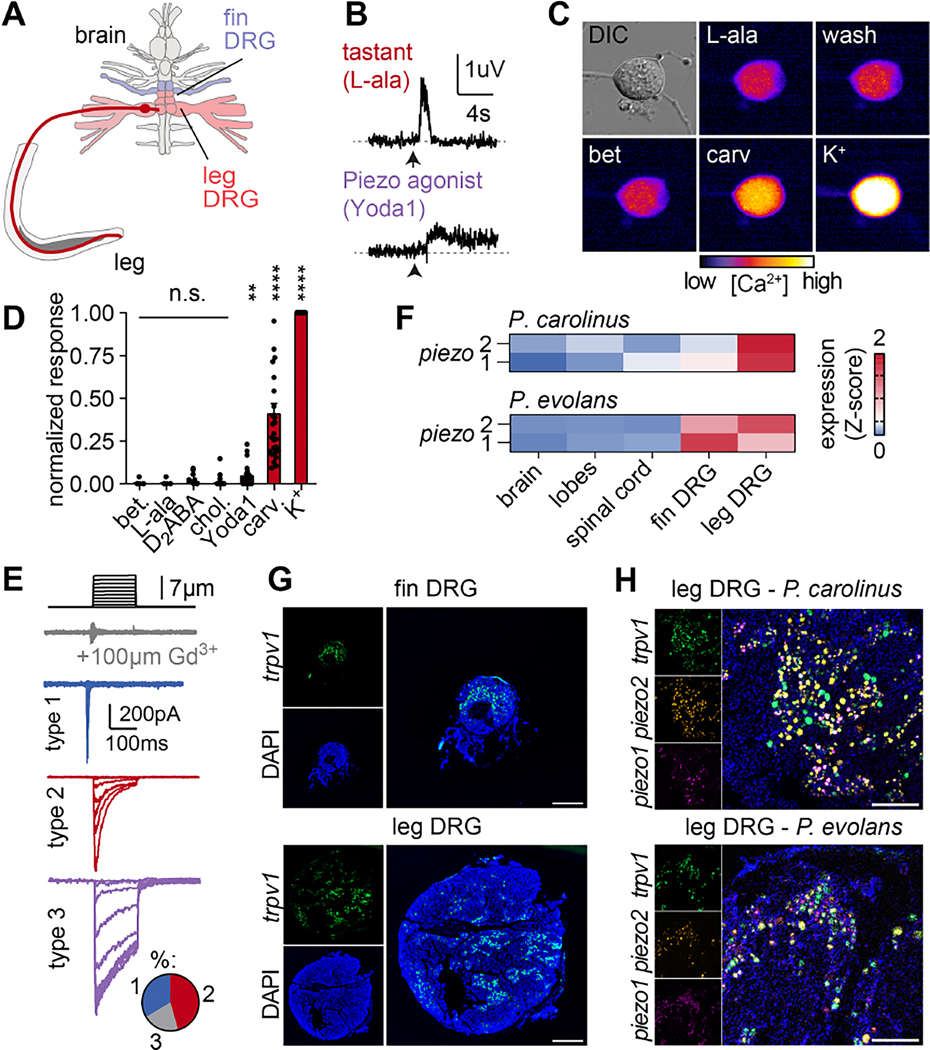
How do animals evolve new traits? Sea robins are fish that possess specialized leg-like appendages used to "walk" along the sea floor. Here, we show that legs are bona fide sense organs that localize buried prey. Legs are covered in sensory papillae that receive dense innervation from touch-sensitive neurons, express non-canonical epithelial taste receptors, and mediate chemical sensitivity that drives predatory digging behavior. A combination of developmental analyses, crosses between species with and without papillae, and interspecies comparisons of sea robins from around the world demonstrate that papillae represent a key evolutionary innovation associated with behavioral niche expansion on the sea floor. These discoveries provide unique insight into how molecular-, cellular-, and tissue-scale adaptations integrate to produce novel organismic traits and behavior.
Keywords: behavioral evolution; chemosensation; comparative biology; evolutionary innovation; leg-like appendages; limb formation; sea robin; sensory papillae; taste receptors; walking fish.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare that they have no competing interests.
Figures


References
-
- Russell M, Grace M, and Gutherz E. (1992). Field Guide to the Searobins (Prionotus and Bellator) in the Western North Atlantic (NOAA)
-
- Bardach JE, and Case J. (1965). Sensory Capabilities of the Modified Fins of Squirrel Hake (Urophycis chuss) and Searobins (Prionotus carolinus and P. evolans). Copeia 1965, 194–206. 10.2307/1440724. - DOI
-
- Manderson JP, Phelan BA, Bejda AJ, Stehlik LL, and Stoner AW (1999). Predation by striped searobin (Prionotus evolans, Triglidae) on young-of-the-year winter flounder (Pseudopleuronectes americanus, Walbaum): examining prey size selection and prey choice using field observations and laboratory experiments. Journal of Experimental Marine Biology and Ecology 242, 211–231. 10.1016/S0022-0981(99)00105-7. - DOI
-
- Ross ST (1977). Patterns of Resource Partitioning in Searobins (Pisces: Triglidae). Copeia 1977, 561–571. 10.2307/1443277. - DOI
-
- Sazima C, and Grossman A. (2005). A non-digging zoobenthivorous fish attracts two opportunistic predatory fish associates. Neotropical Ichthyology 3, 445–448. 10.1590/S1679-62252005000300014. - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
