

Phosphoproteomics-directed manipulation reveals SEC22B as a hepatocellular signaling node governing metabolic actions of glucagon
- PMID: 39333498
- PMCID: PMC11436942
- DOI: 10.1038/s41467-024-52703-w
Phosphoproteomics-directed manipulation reveals SEC22B as a hepatocellular signaling node governing metabolic actions of glucagon
Abstract
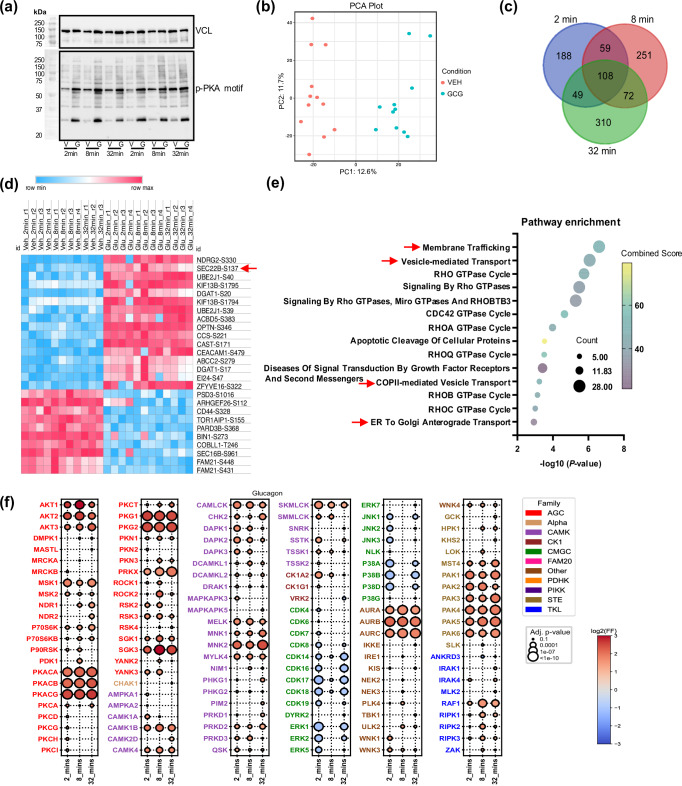
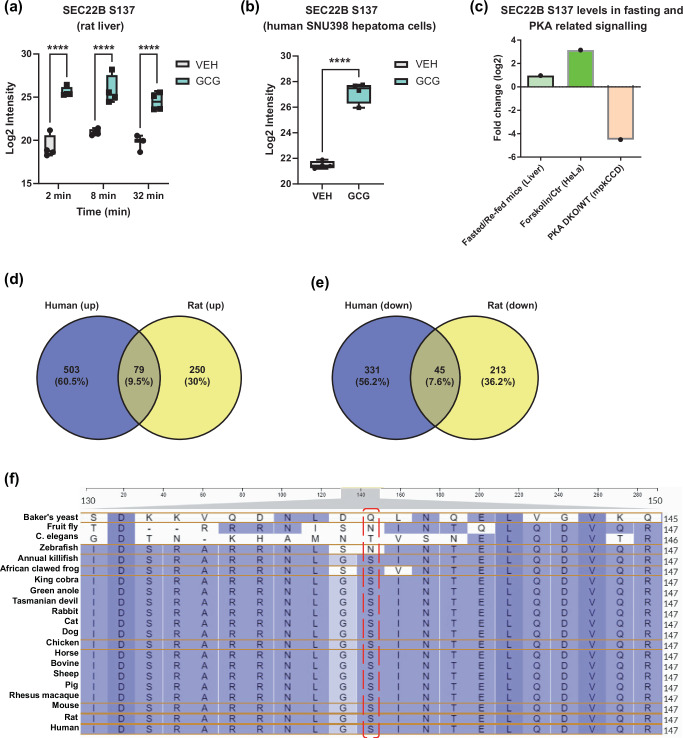
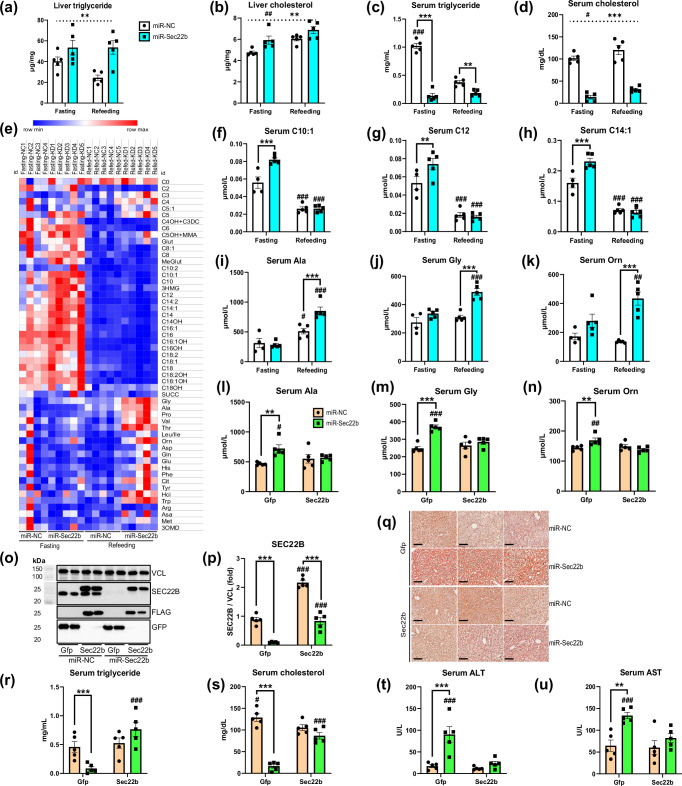
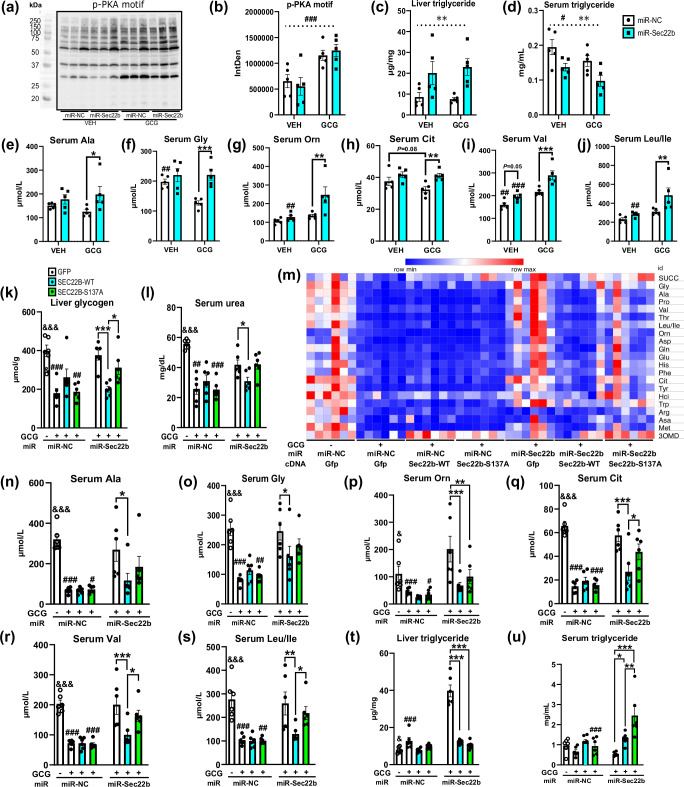
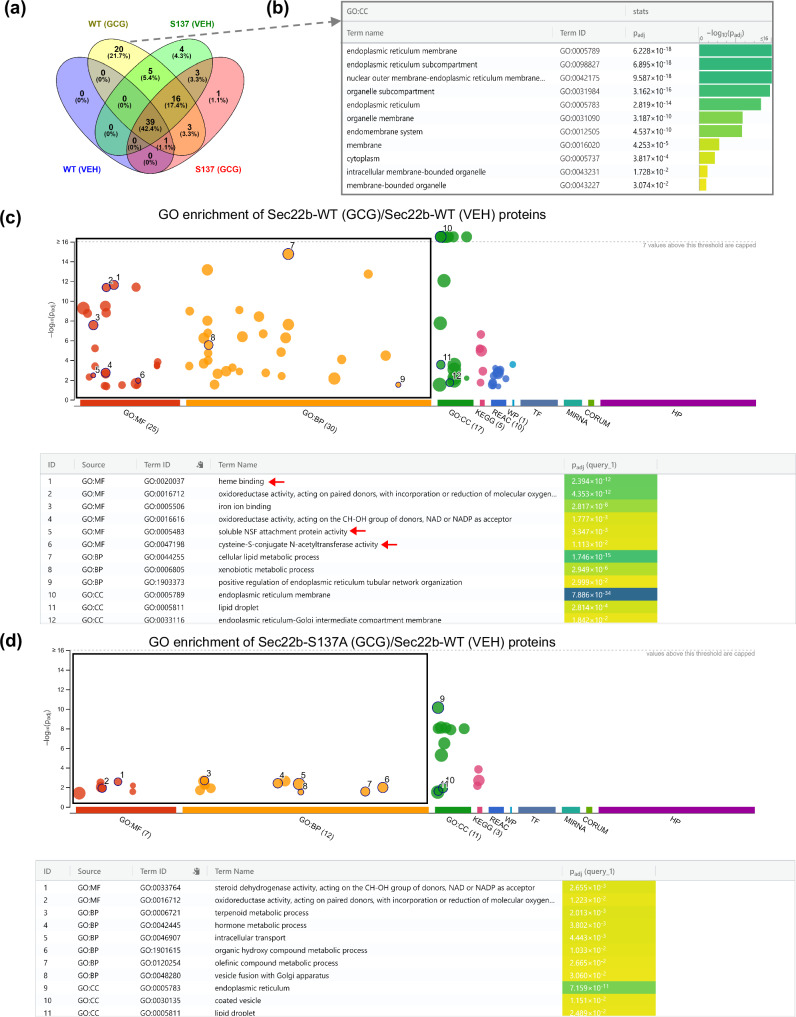
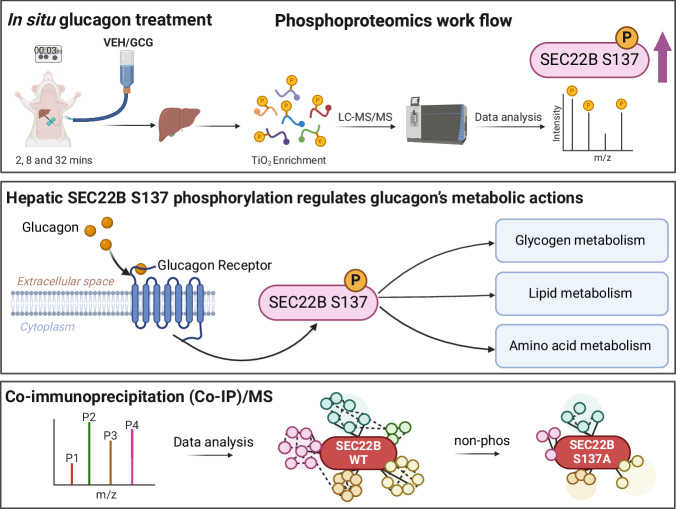
The peptide hormone glucagon is a fundamental metabolic regulator that is also being considered as a pharmacotherapeutic option for obesity and type 2 diabetes. Despite this, we know very little regarding how glucagon exerts its pleiotropic metabolic actions. Given that the liver is a chief site of action, we performed in situ time-resolved liver phosphoproteomics to reveal glucagon signaling nodes. Through pathway analysis of the thousands of phosphopeptides identified, we reveal "membrane trafficking" as a dominant signature with the vesicle trafficking protein SEC22 Homolog B (SEC22B) S137 phosphorylation being a top hit. Hepatocyte-specific loss- and gain-of-function experiments reveal that SEC22B was a key regulator of glycogen, lipid and amino acid metabolism, with SEC22B-S137 phosphorylation playing a major role in glucagon action. Mechanistically, we identify several protein binding partners of SEC22B affected by glucagon, some of which were differentially enriched with SEC22B-S137 phosphorylation. In summary, we demonstrate that phosphorylation of SEC22B is a hepatocellular signaling node mediating the metabolic actions of glucagon and provide a rich resource for future investigations on the biology of glucagon action.
© 2024. The Author(s).
Conflict of interest statement
A.J.R and P.M.R receive research support from Boehringer-Ingelheim, a pharmaceutical company, for an unrelated research project. L.C.C is a founder and member of the board of directors of Agios Pharmaceuticals and is a founder and receives research support from Petra Pharmaceuticals; is listed as an inventor on a patent (WO2019232403A1, Weill Cornell Medicine) for combination therapy for PI3K-associated disease or disorder, and the identification of therapeutic interventions to improve response to PI3K inhibitors for cancer treatment; is a co-founder and shareholder in Faeth Therapeutics; has equity in and consults for Cell Signaling Technologies, Volastra, Larkspur, Omega Funds and 1 Base Pharmaceuticals; and consults for Loxo-Lilly. T.M.Y. is a co-founder of DeStroke. J.L.J has received consulting fees from Scorpion Therapeutics and Volastra Therapeutics. All other authors declare no conflicts of interest.
Figures
References
-
- Kimball C. P. & Murlin J. R. Aqueous extracts of pancreas: III. Some precipitation reactions of insulin. J. Biol. Chem.58, (1923).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
