

Lipid droplets sequester palmitic acid to disrupt endothelial ciliation and exacerbate atherosclerosis in male mice
- PMID: 39333556
- PMCID: PMC11437155
- DOI: 10.1038/s41467-024-52621-x
Lipid droplets sequester palmitic acid to disrupt endothelial ciliation and exacerbate atherosclerosis in male mice
Abstract
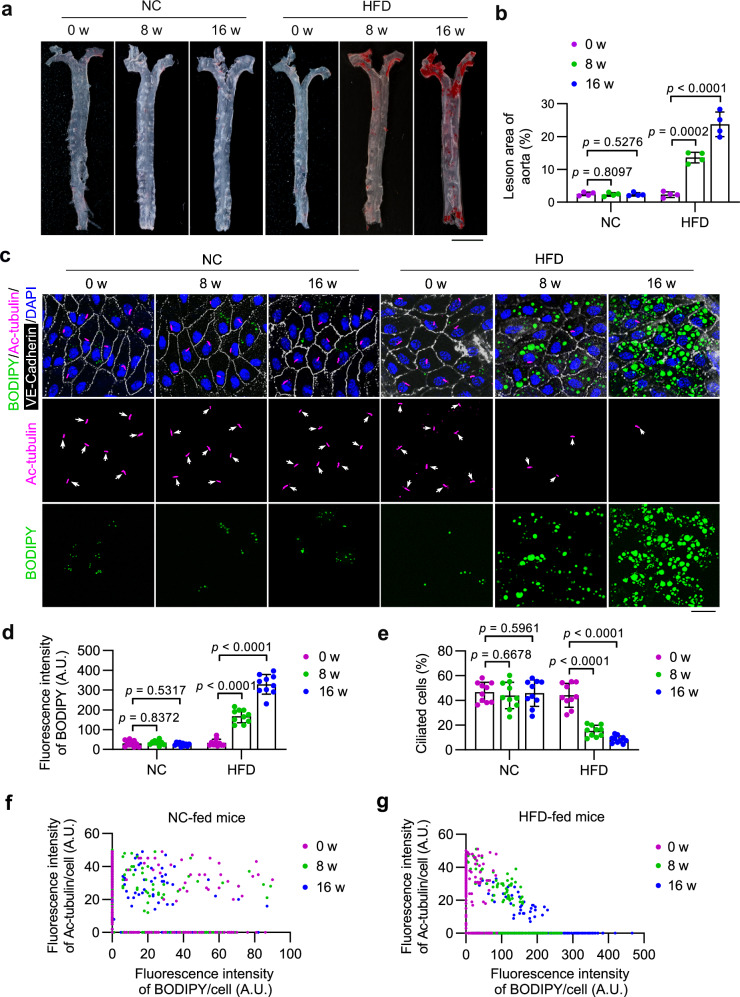
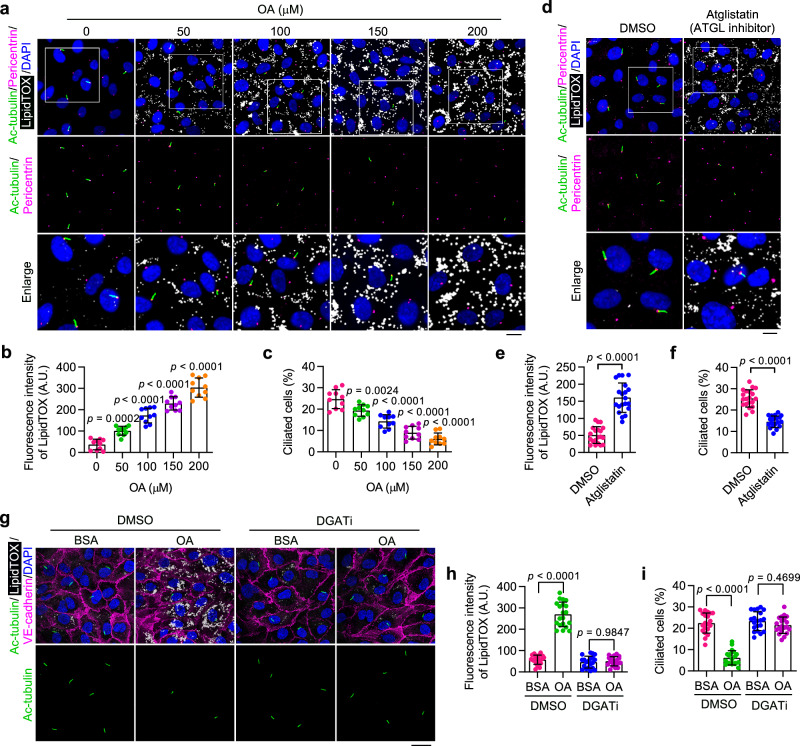
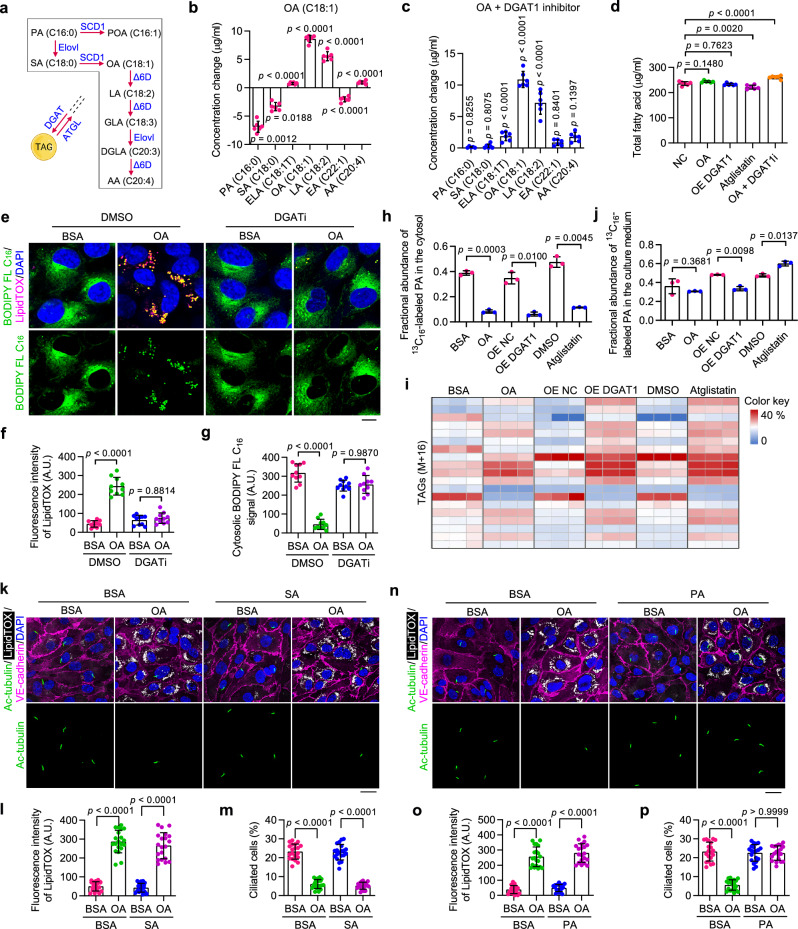
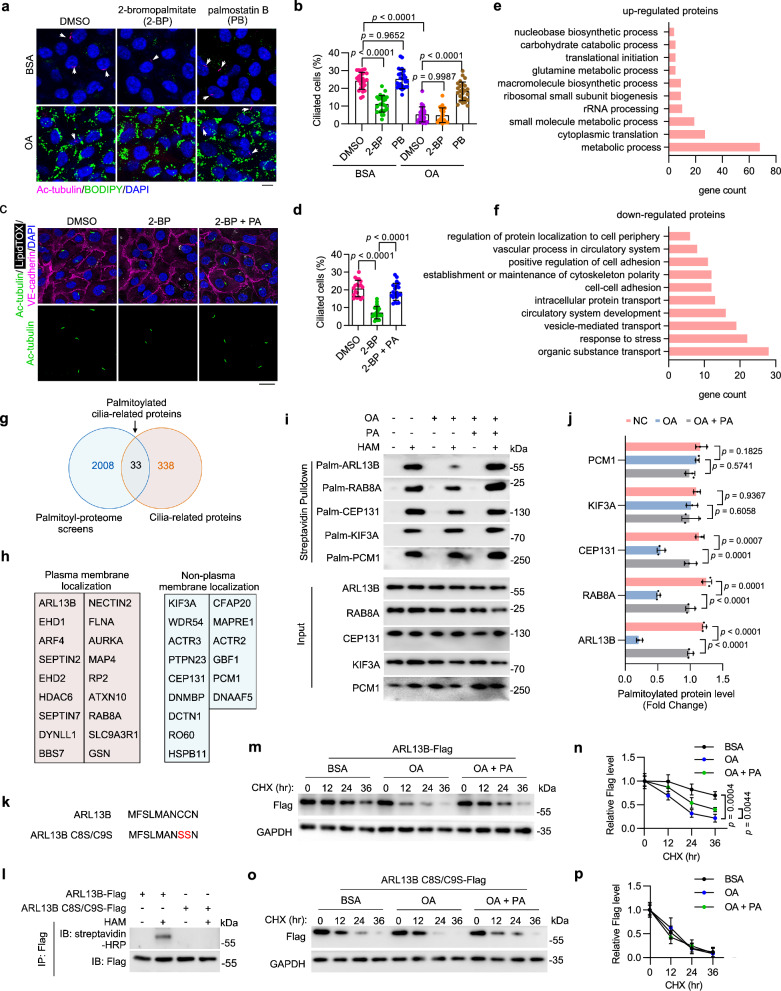
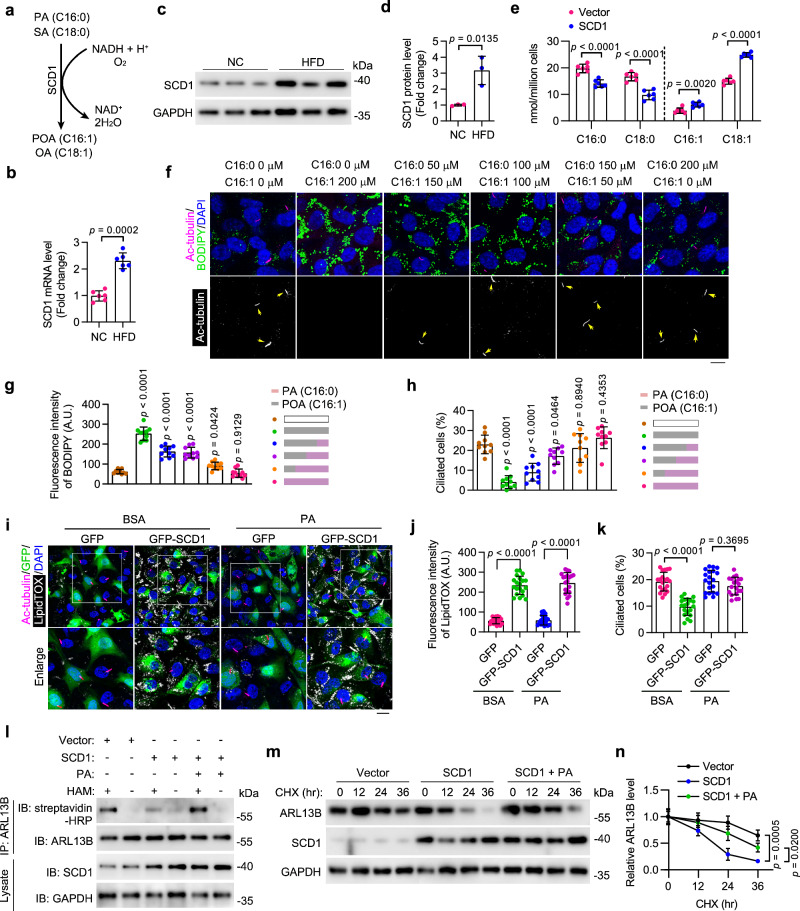
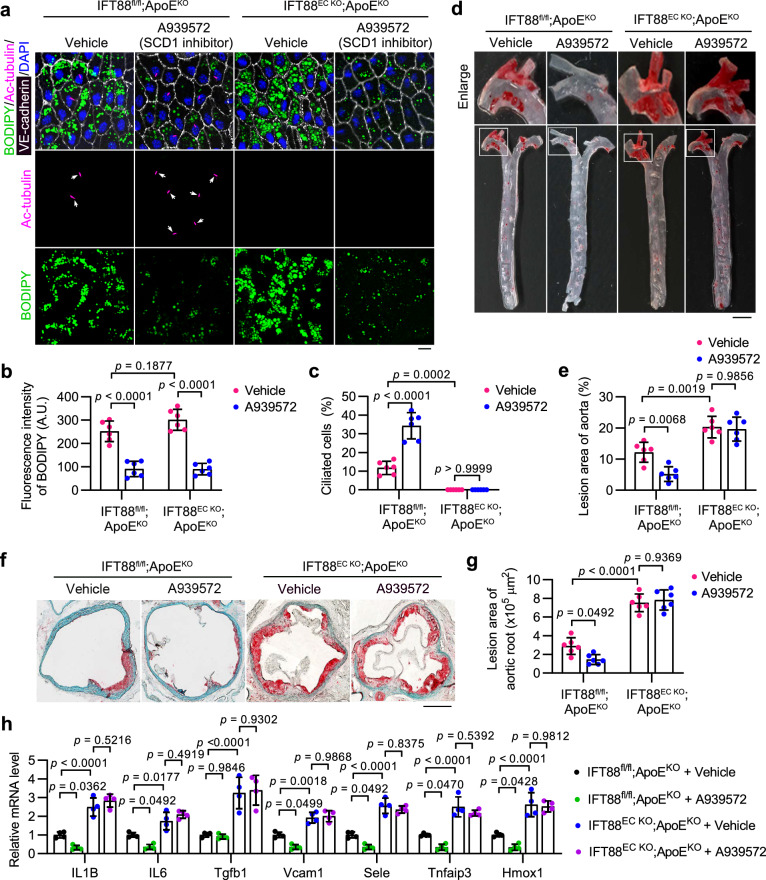
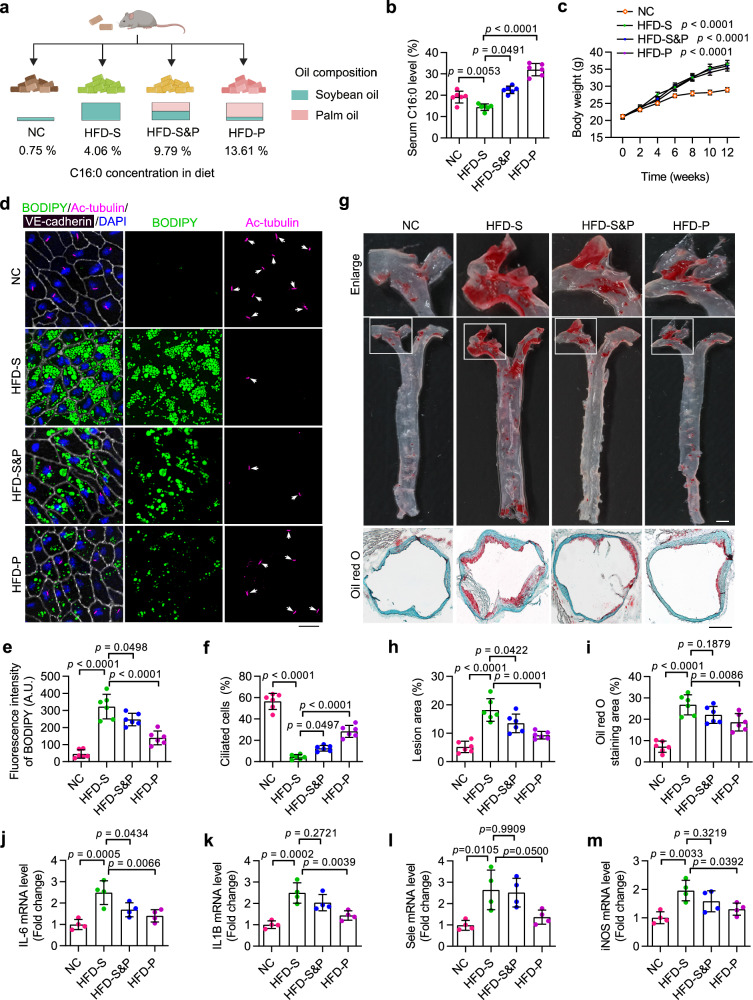
Disruption of ciliary homeostasis in vascular endothelial cells has been implicated in the development of atherosclerosis. However, the molecular basis for the regulation of endothelial cilia during atherosclerosis remains poorly understood. Herein, we provide evidence in male mice that the accumulation of lipid droplets in vascular endothelial cells induces ciliary loss and contributes to atherosclerosis. Triglyceride accumulation in vascular endothelial cells differentially affects the abundance of free fatty acid species in the cytosol, leading to stimulated lipid droplet formation and suppressed protein S-palmitoylation. Reduced S-palmitoylation of ciliary proteins, including ADP ribosylation factor like GTPase 13B, results in the loss of cilia. Restoring palmitic acid availability, either through pharmacological inhibition of stearoyl-CoA desaturase 1 or a palmitic acid-enriched diet, significantly restores endothelial cilia and mitigates the progression of atherosclerosis. These findings thus uncover a previously unrecognized role of lipid droplets in regulating ciliary homeostasis and provide a feasible intervention strategy for preventing and treating atherosclerosis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Liu, M. et al. Macrophage K63-linked ubiquitination of YAP promotes its nuclear localization and exacerbates atherosclerosis. Cell Rep.32, 107990 (2020). - PubMed
-
- Karunakaran, D. et al. RIPK1 expression associates with inflammation in early atherosclerosis in humans and can be therapeutically silenced to reduce NF-κB activation and atherogenesis in mice. Circulation143, 163–177 (2021). - PubMed
-
- Jia, M. et al. Deletion of BACH1 attenuates atherosclerosis by reducing endothelial inflammation. Circ. Res.130, 1038–1055 (2022). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
