

Modulation of cannabinoid receptor 2 alters neuroinflammation and reduces formation of alpha-synuclein aggregates in a rat model of nigral synucleinopathy
- PMID: 39334169
- PMCID: PMC11438102
- DOI: 10.1186/s12974-024-03221-5
Modulation of cannabinoid receptor 2 alters neuroinflammation and reduces formation of alpha-synuclein aggregates in a rat model of nigral synucleinopathy
Abstract
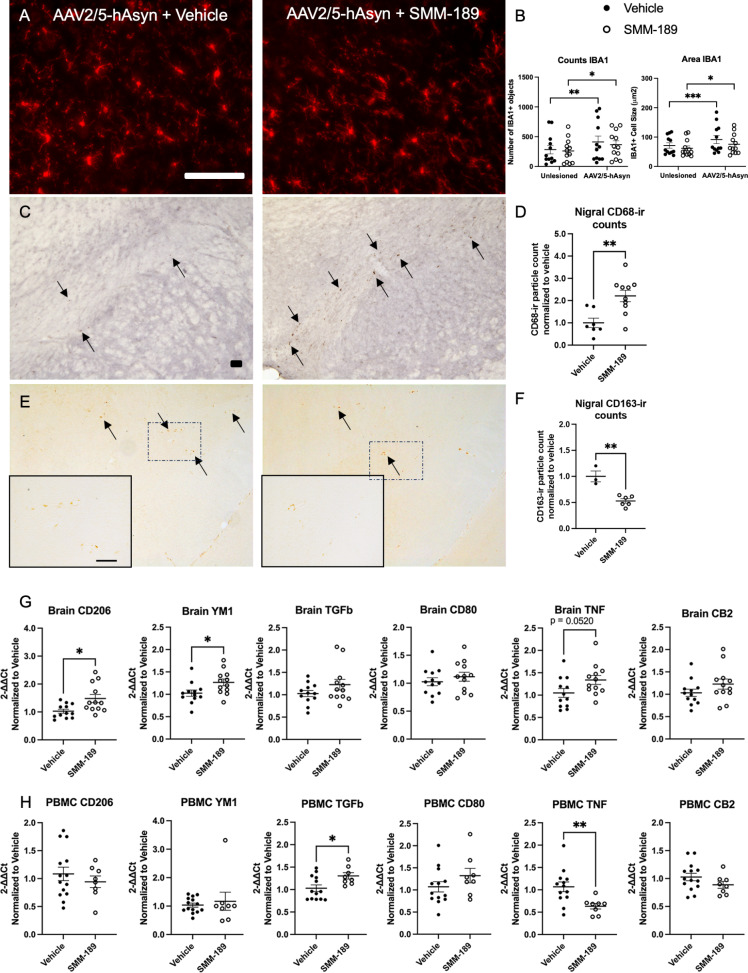
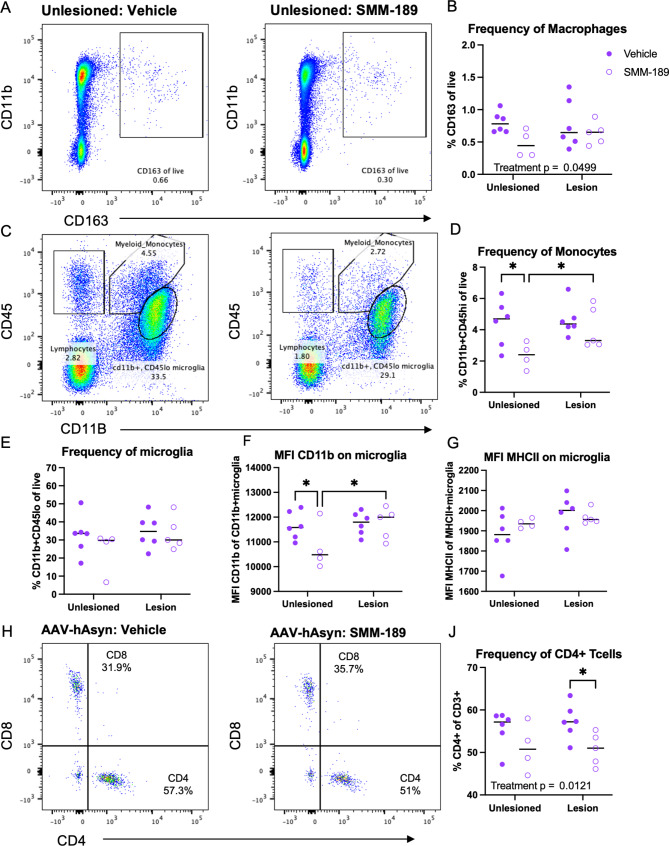
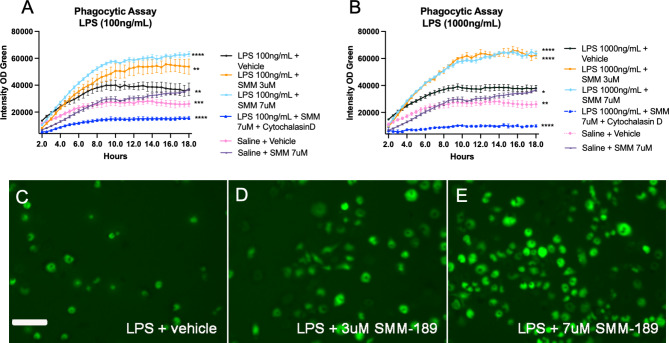
Research into the disequilibrium of microglial phenotypes has become an area of intense focus in neurodegenerative disease as a potential mechanism that contributes to chronic neuroinflammation and neuronal loss in Parkinson's disease (PD). There is growing evidence that neuroinflammation accompanies and may promote progression of alpha-synuclein (Asyn)-induced nigral dopaminergic (DA) degeneration. From a therapeutic perspective, development of immunomodulatory strategies that dampen overproduction of pro-inflammatory cytokines from chronically activated immune cells and induce a pro-phagocytic phenotype is expected to promote Asyn removal and protect vulnerable neurons. Cannabinoid receptor-2 (CB2) is highly expressed on activated microglia and peripheral immune cells, is upregulated in the substantia nigra of individuals with PD and in mouse models of nigral degeneration. Furthermore, modulation of CB2 protects against rotenone-induced nigral degeneration; however, CB2 has not been pharmacologically and selectively targeted in an Asyn model of PD. Here, we report that 7 weeks of peripheral administration of CB2 inverse agonist SMM-189 reduced phosphorylated (pSer129) Asyn in the substantia nigra compared to vehicle treatment. Additionally, SMM-189 delayed Asyn-induced immune cell infiltration into the brain as determined by flow cytometry, increased CD68 protein expression, and elevated wound-healing-immune-mediator gene expression. Additionally, peripheral immune cells increased wound-healing non-classical monocytes and decreased pro-inflammatory classical monocytes. In vitro analysis of RAW264.7 macrophages treated with lipopolysaccharide (LPS) and SMM-189 revealed increased phagocytosis as measured by the uptake of fluorescence of pHrodo E. coli bioparticles. Together, results suggest that targeting CB2 with SMM-189 skews immune cell function toward a phagocytic phenotype and reduces toxic aggregated species of Asyn. Our novel findings demonstrate that CB2 may be a target to modulate inflammatory and immune responses in proteinopathies.
Keywords: Alpha-synuclein; Cannabinoid receptor-2; Microglia phenotype; Parkinson’s disease.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Update of
-
Modulation of cannabinoid receptor 2 alters neuroinflammation and reduces formation of alpha-synuclein aggregates in a rat model of nigral synucleinopathy.bioRxiv [Preprint]. 2024 Mar 23:2023.08.25.554814. doi: 10.1101/2023.08.25.554814. bioRxiv. 2024. Update in: J Neuroinflammation. 2024 Sep 27;21(1):240. doi: 10.1186/s12974-024-03221-5. PMID: 38562842 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
