

Analysis of early effects of human APOE isoforms on Alzheimer's disease and type III hyperlipoproteinemia pathways using knock-in rat models with humanized APP and APOE
- PMID: 39334477
- PMCID: PMC11438110
- DOI: 10.1186/s12964-024-01832-2
Analysis of early effects of human APOE isoforms on Alzheimer's disease and type III hyperlipoproteinemia pathways using knock-in rat models with humanized APP and APOE
Erratum in
-
Correction: Analysis of early effects of human APOE isoforms on Alzheimer's disease and type III hyperlipoproteinemia pathways using knock-in rat models with humanized APP and APOE.Cell Commun Signal. 2024 Dec 31;22(1):619. doi: 10.1186/s12964-024-02004-y. Cell Commun Signal. 2024. PMID: 39741282 Free PMC article. No abstract available.
Abstract
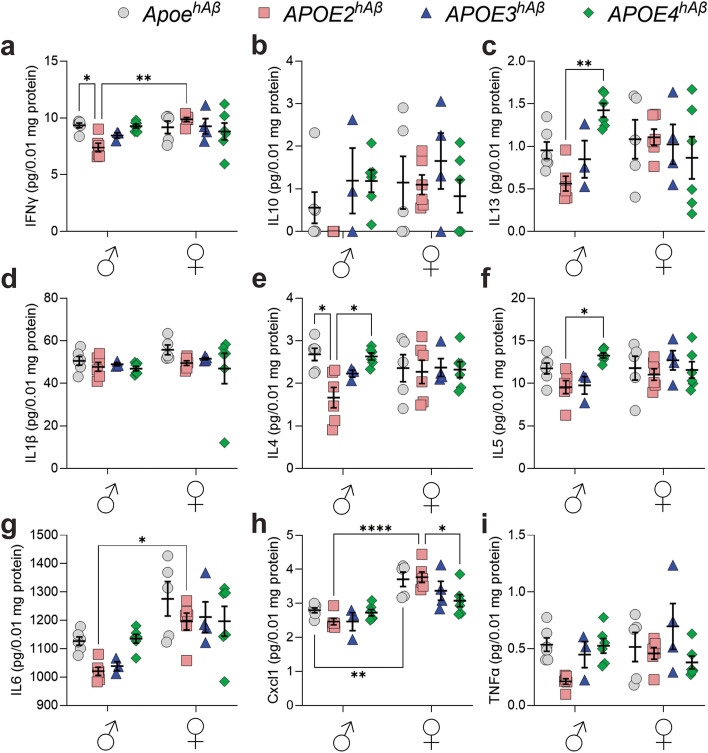
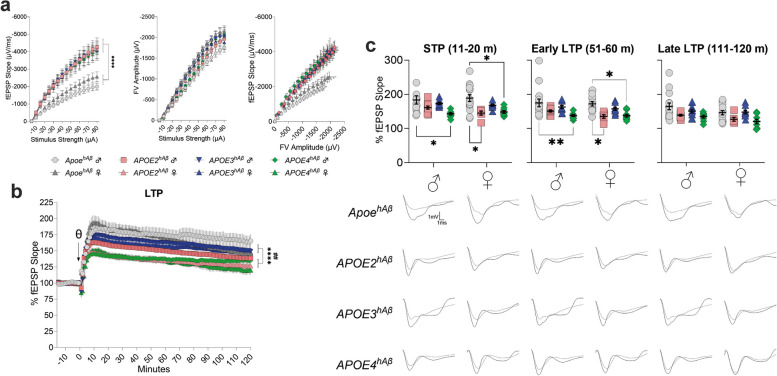
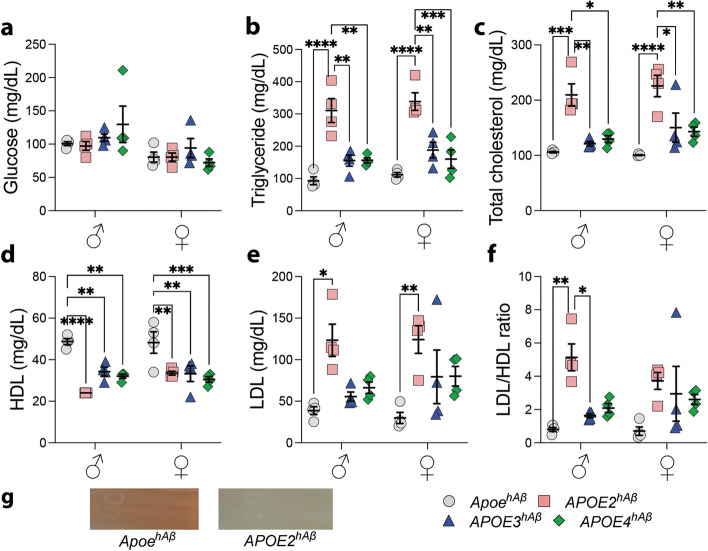
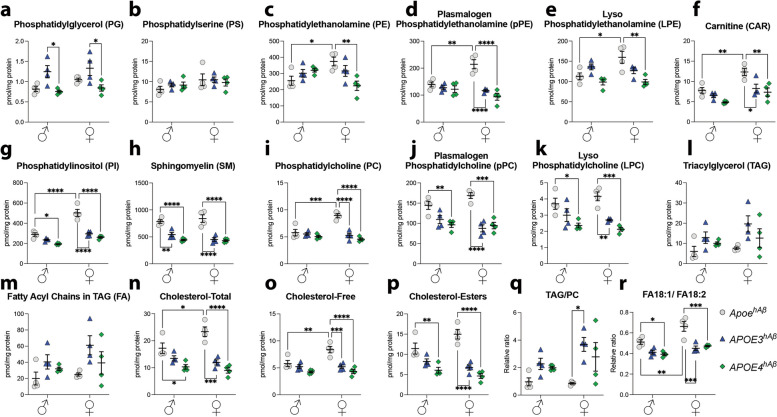
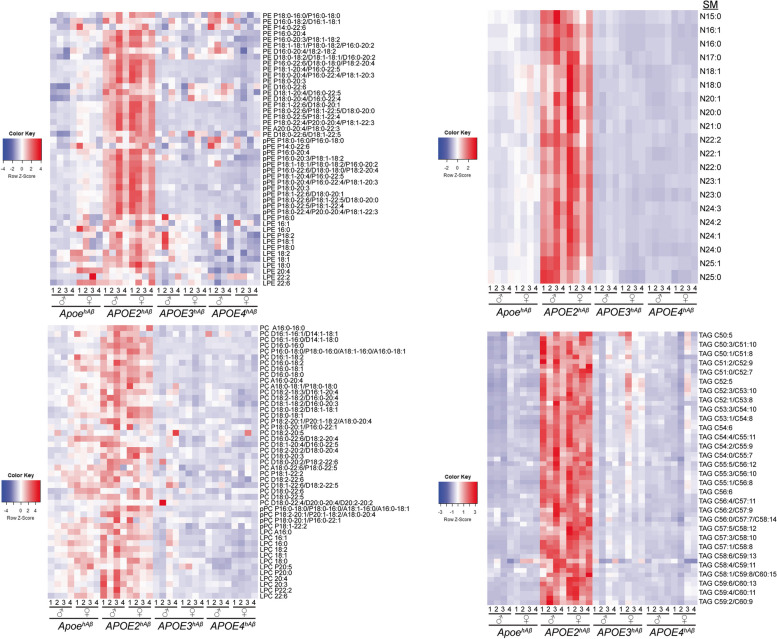
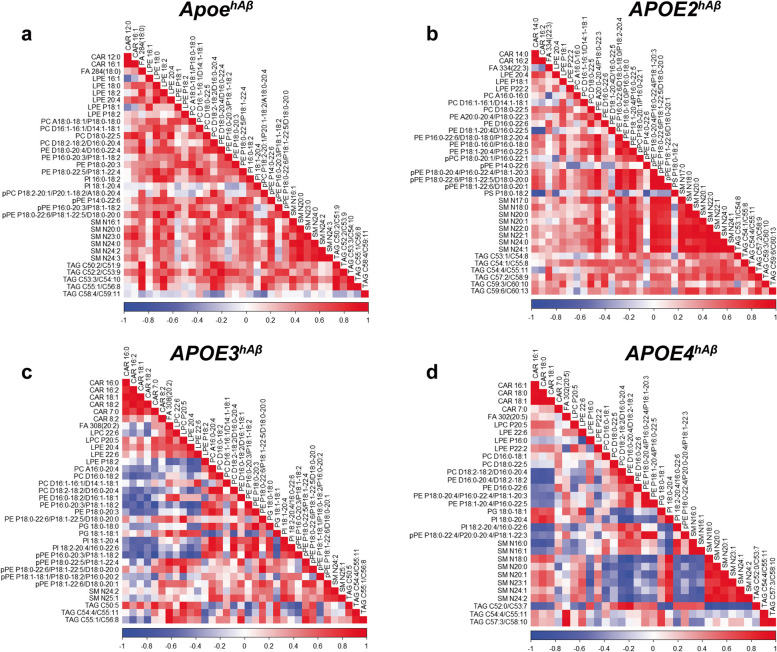
APOE is a major genetic factor in late-onset Alzheimer's disease (LOAD), with APOE4 increasing risk, APOE3 acting as neutral, and APOE2 offering protection. APOE also plays key role in lipid metabolism, affecting both peripheral and central systems, particularly in lipoprotein metabolism in triglyceride and cholesterol regulation. APOE2 is linked to Hyperlipoproteinemia type III (HLP), characterized by mixed hypercholesterolemia and hypertriglyceridemia due to impaired binding to Low-Density Lipoproteins receptors. To explore the impact of human APOE isoforms on LOAD and lipid metabolism, we developed Long-Evans rats with human APOE2, APOE3, or APOE4 in place of rat Apoe. These rats were crossed with those carrying a humanized App allele to express human Aβ, which is more aggregation-prone than rodent Aβ, enabling the study of human APOE-human Aβ interactions. In this study, we focused on 80-day-old adolescent rats to analyze early changes that may be associated with the later development of LOAD. We found that APOE2hAβ rats had the highest levels of APOE in serum and brain, with no significant transcriptional differences among isoforms, suggesting variations in protein translation or stability. Aβ43 levels were significantly higher in male APOE4hAβ rats compared to APOE2hAβ rats. However, no differences in Tau or phosphorylated Tau levels were observed across the APOE isoforms. Neuroinflammation analysis revealed lower levels of IL13, IL4 and IL5 in APOE2hAβ males compared to APOE4hAβ males. Neuronal transmission and plasticity tests using field Input-Output (I/O) and long-term potentiation (LTP) recordings showed increased excitability in all APOE-carrying rats, with LTP deficits in APOE2hAβand APOE4hAβ rats compared to ApoehAβ and APOE3hAβ rats. Additionally, a lipidomic analysis of 222 lipid molecular species in serum samples showed that APOE2hAβ rats displayed elevated triglycerides and cholesterol, making them a valuable model for studying HLP. These rats also exhibited elevated levels of phosphatidylglycerol, phosphatidylserine, phosphatidylethanolamine, sphingomyelin, and lysophosphatidylcholine. Minimal differences in lipid profiles between APOE3hAβ and APOE4hAβ rats reflect findings from mouse models. Future studies will include comprehensive lipidomic analyses in various CNS regions and at older ages to further validate these models and explore the effects of APOE isoforms on lipid metabolism in relation to AD pathology.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
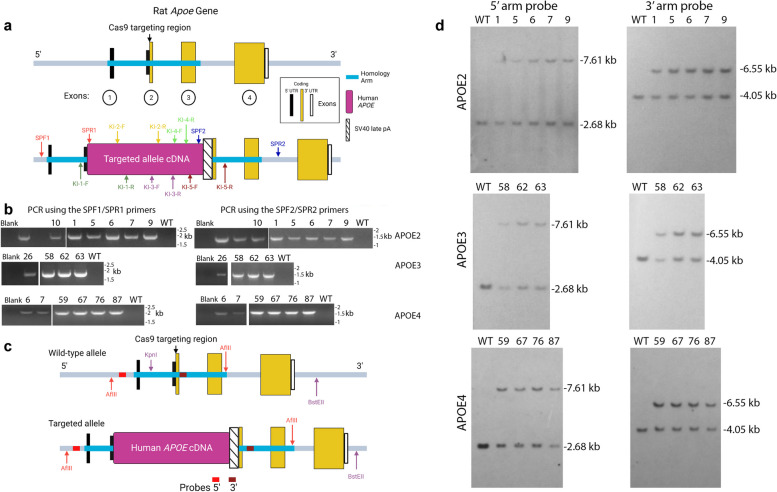
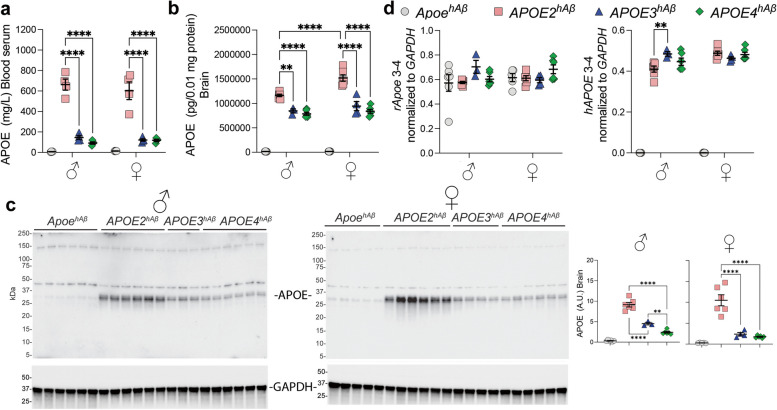
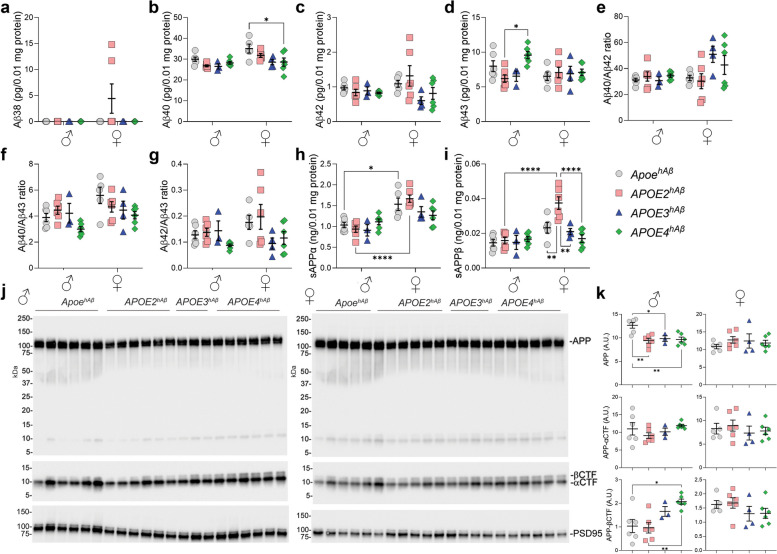
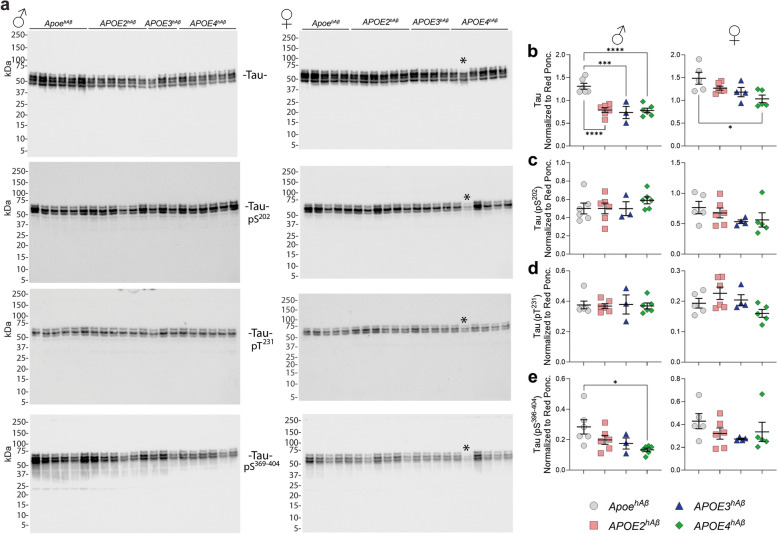


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
