

A Quadruple Gene-Deleted Live BoHV-1 Subunit RVFV Vaccine Vector Reactivates from Latency and Replicates in the TG Neurons of Calves but Is Not Transported to and Shed from Nasal Mucosa
- PMID: 39339973
- PMCID: PMC11437494
- DOI: 10.3390/v16091497
A Quadruple Gene-Deleted Live BoHV-1 Subunit RVFV Vaccine Vector Reactivates from Latency and Replicates in the TG Neurons of Calves but Is Not Transported to and Shed from Nasal Mucosa
Abstract
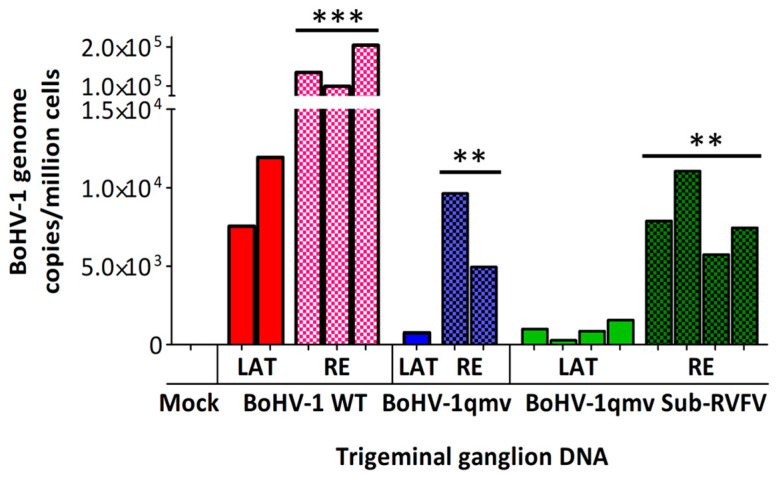
Bovine herpesvirus type 1 (BoHV-1) establishes lifelong latency in trigeminal ganglionic (TG) neurons following intranasal and ocular infection in cattle. Periodically, the latent virus reactivates in the TG due to stress and is transported anterogradely to nerve endings in the nasal epithelium, where the virus replicates and sheds. Consequently, BoHV-1 is transmitted to susceptible animals and maintained in the cattle population. Modified live BoHV-1 vaccine strains (BoHV-1 MLV) also have a similar latency reactivation. Therefore, they circulate and are maintained in cattle herds. Additionally, they can regain virulence and cause vaccine outbreaks because they mutate and recombine with other circulating field wild-type (wt) strains. Recently, we constructed a BoHV-1 quadruple mutant virus (BoHV-1qmv) that lacks immune evasive properties due to UL49.5 and glycoprotein G (gG) deletions. In addition, it also lacks the gE cytoplasmic tail (gE CT) and Us9 gene sequences designed to make it safe, increase its vaccine efficacy against BoHV-1, and restrict its anterograde neuronal transport noted above. Further, we engineered the BoHV-1qmv-vector to serve as a subunit vaccine against the Rift Valley fever virus (BoHV-1qmv Sub-RVFV) (doi: 10.3390/v15112183). In this study, we determined the latency reactivation and nasal virus shedding properties of BoHV-1qmv (vector) and BoHV-1qmv-vectored subunit RVFV (BoHV-1qmv sub-RVFV) vaccine virus in calves in comparison to the BoHV-1 wild-type (wt) following intranasal inoculation. The real-time PCR results showed that BoHV-1 wt- but not the BoHV-1qmv vector- and BoHV-1qmv Sub-RVFV-inoculated calves shed virus in the nose following dexamethasone-induced latency reactivation; however, like the BoHV-1 wt, both the BoHV-1qmv vector and BoHV-1qmv Sub-RVFV viruses established latency, were reactivated, and replicated in the TG neurons. These results are consistent with the anterograde neurotransport function of the gE CT and Us9 sequences, which are deleted in the BoHV-1qmv and BoHV-1qmv Sub-RVFV.
Keywords: BoHV-1; BoHV-1qmv vector; DIVA; bovine herpesvirus; cattle; glycoprotein E; latency reactivation; quadruple mutant virus; trigeminal ganglion; vectored vaccine.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Zhou Y., Shao Z., Dai G., Li X., Xiang Y., Jiang S., Zhang Z., Ren Y., Zhu Z., Fan C., et al. Pathogenic infection characteristics and risk factors for bovine respiratory disease complex based on the detection of lung pathogens in dead cattle in Northeast China. J. Dairy. Sci. 2023;106:589–606. doi: 10.3168/jds.2022-21929. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
