

A Single-Component Multilayered Self-Assembling Protein Nanoparticle Vaccine Based on Extracellular Domains of Matrix Protein 2 against Both Influenza A and B
- PMID: 39340007
- PMCID: PMC11435909
- DOI: 10.3390/vaccines12090975
A Single-Component Multilayered Self-Assembling Protein Nanoparticle Vaccine Based on Extracellular Domains of Matrix Protein 2 against Both Influenza A and B
Abstract
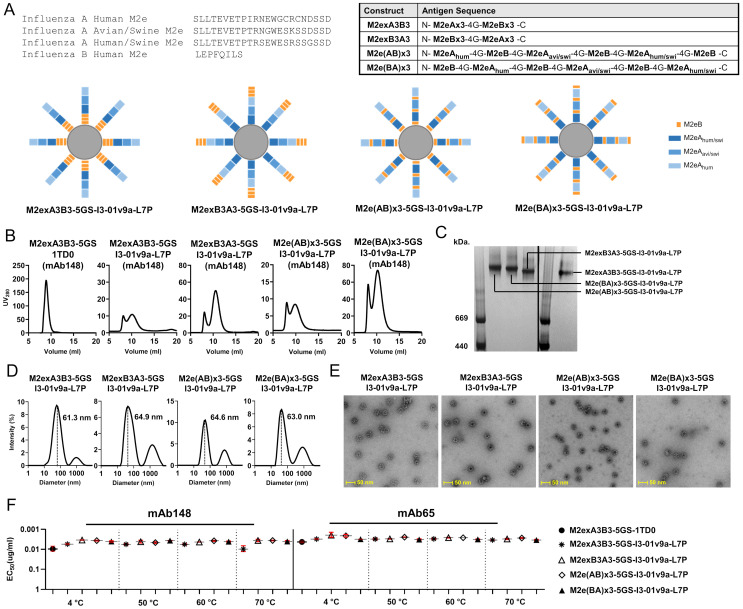
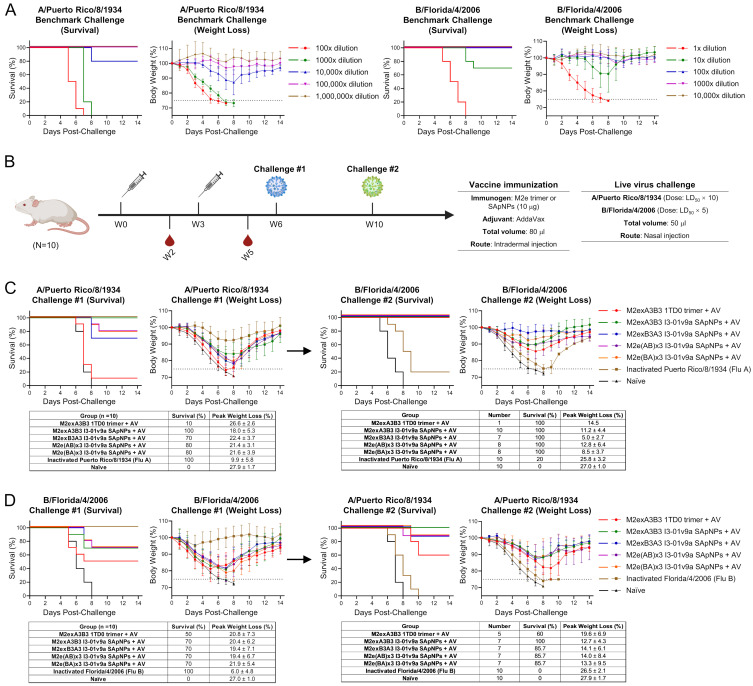
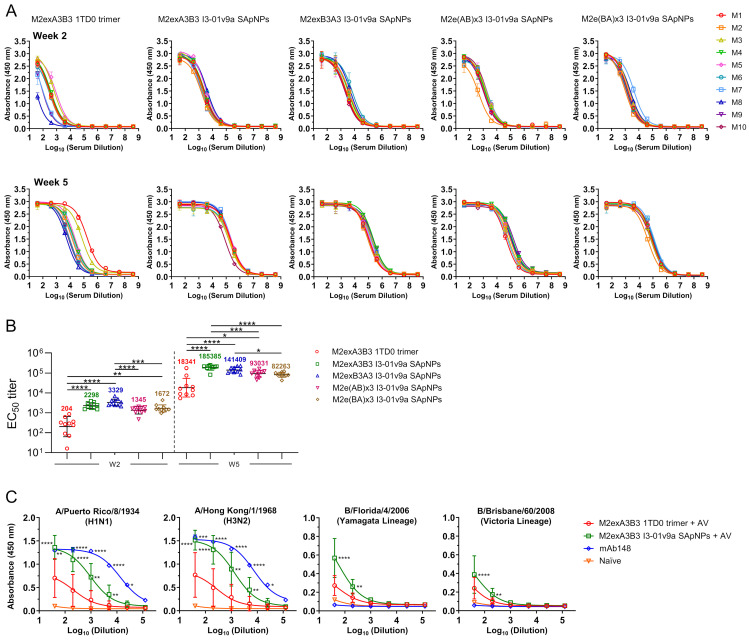
The development of an effective and broadly protective influenza vaccine against circulating and emerging strains remains elusive. In this study, we evaluated a potentially universal influenza vaccine based on single-component self-assembling protein nanoparticles (1c-SApNPs) presenting the conserved matrix protein 2 ectodomain (M2e) from influenza A and B viruses (IAV and IBV, respectively). We previously designed a tandem antigen comprising three IAV M2e domains of human, avian/swine, and human/swine origins (termed M2ex3). The M2ex3-presenting 1c-SApNPs conferred complete protection in mice against sequential lethal challenges with H1N1 and H3N2. To broaden this protection to cover IBVs, we designed a series of antigens incorporating different arrangements of three IAV M2e domains and three copies of IBV M2e. Tandem repeats of IAV and IBV (termed influenza A-B) M2e arrayed on the I3-01v9a 60-mer 1c-SApNP, when formulated with an oil-in-water emulsion adjuvant, generated greater M2e-specific immunogenicity and protective efficacy than the soluble influenza A-B M2e trimer, indicated by higher survival rates and reduced weight loss post-challenge. Importantly, one of the influenza A-B M2e SApNP constructs elicited 100% protection against a lethal influenza A/Puerto Rico/8/1934 (H1N1) challenge in mice and 70% protection against a lethal influenza B/Florida/4/2006 (Yamagata lineage) challenge, the latter of which has not been reported in the literature to date. Our study thus provides a promising M2e-based single-component universal vaccine candidate against the two major types of influenza virus circulating in humans.
Keywords: M2e; antibody; influenza A; influenza B; protein nanoparticle; universal influenza vaccine.
Conflict of interest statement
Jiang Zhu serves as the Co-Founder, Interim Chief Scientific Officer, Consultant, and Scientific Advisory Board member of Uvax Bio, LLC, and holds associated financial interests. Other authors declare that they have no competing interests.
Figures
Similar articles
-
Single-Component Multilayered Self-Assembling Protein Nanoparticles Displaying Extracellular Domains of Matrix Protein 2 as a Pan-influenza A Vaccine.ACS Nano. 2023 Dec 12;17(23):23545-23567. doi: 10.1021/acsnano.3c06526. Epub 2023 Nov 21. ACS Nano. 2023. PMID: 37988765 Free PMC article.
-
An experimental universal swine influenza a virus (IAV) vaccine candidate based on the M2 ectodomain (M2e) peptide does not provide protection against H1N1 IAV challenge in pigs.Vaccine. 2024 Jan 12;42(2):220-228. doi: 10.1016/j.vaccine.2023.12.012. Epub 2023 Dec 11. Vaccine. 2024. PMID: 38087714
-
Improving Cross-Protection against Influenza Virus Using Recombinant Vaccinia Vaccine Expressing NP and M2 Ectodomain Tandem Repeats.Virol Sin. 2019 Oct;34(5):583-591. doi: 10.1007/s12250-019-00138-9. Epub 2019 Jun 25. Virol Sin. 2019. PMID: 31240620 Free PMC article.
-
M2e-based universal influenza A vaccine.Vaccine. 2009 Oct 23;27(45):6280-3. doi: 10.1016/j.vaccine.2009.07.007. Vaccine. 2009. PMID: 19840661 Review.
-
M2e-Based Influenza Vaccines with Nucleoprotein: A Review.Vaccines (Basel). 2021 Jul 4;9(7):739. doi: 10.3390/vaccines9070739. Vaccines (Basel). 2021. PMID: 34358155 Free PMC article. Review.
Cited by
-
Rational design of next-generation filovirus vaccines with glycoprotein stabilization, nanoparticle display, and glycan modification.bioRxiv [Preprint]. 2025 Mar 2:2025.03.02.641072. doi: 10.1101/2025.03.02.641072. bioRxiv. 2025. PMID: 40060701 Free PMC article. Preprint.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
