

c-CBL/LCK/c-JUN/ETS1/CD28 axis restrains childhood asthma by suppressing Th2 differentiation
- PMID: 39342146
- PMCID: PMC11439220
- DOI: 10.1186/s10020-024-00872-1
c-CBL/LCK/c-JUN/ETS1/CD28 axis restrains childhood asthma by suppressing Th2 differentiation
Abstract
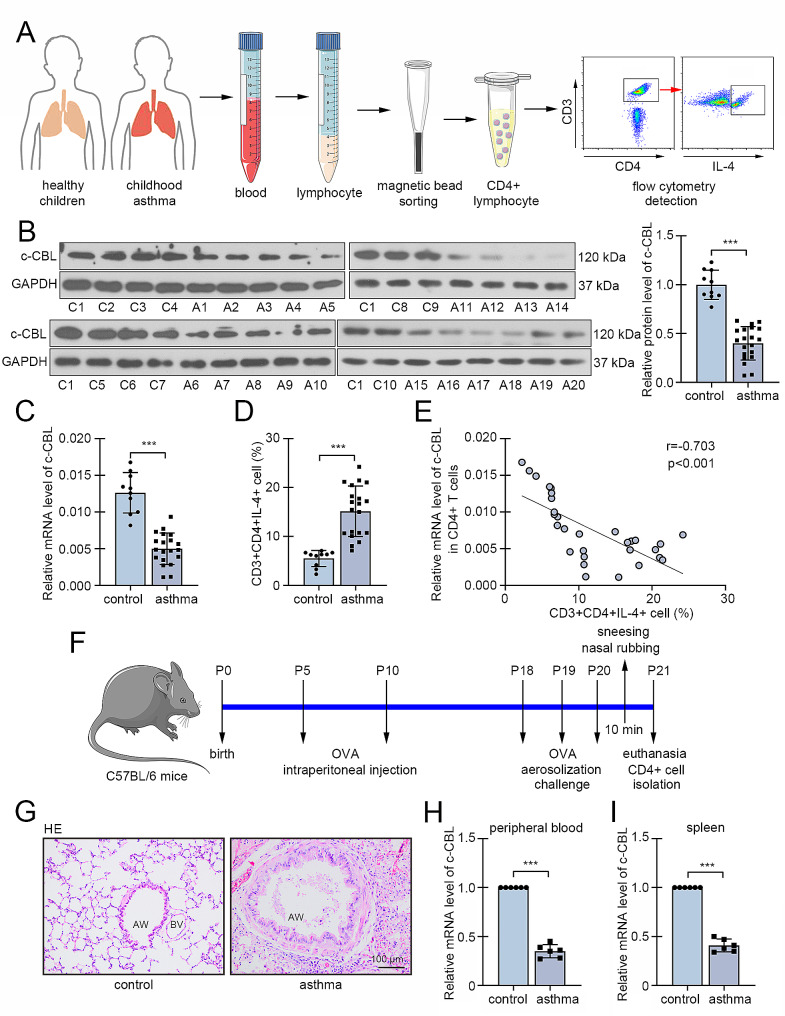
Background: Asthma is a common immune disease with high morbidity in children. Type 2 inflammation is the center of asthma development, and mainly mediated by a subset of CD4 + T cells, T helper 2 (Th2) cells. Excess Th2 differentiation was generally associated with asthmatic attack. Casitas B-lineage lymphoma (c-CBL) was reported to involved in T cell development and databank showed its decreased expression in CD4 + T cells from peripheral blood of asthmatic children. This study aims to investigate the role of c-CBL in childhood asthma and Th2 differentiation, and explore the underlying mechanism.
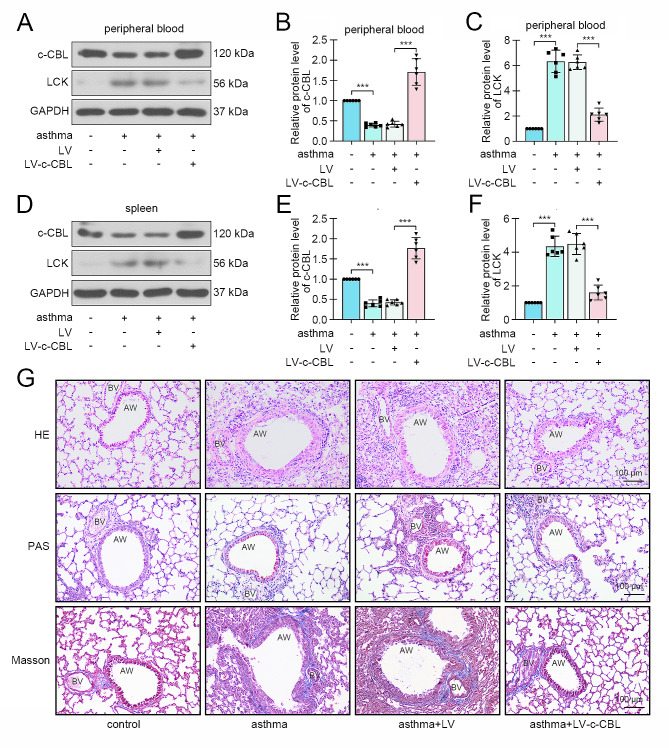
Methods: We collected peripheral blood samples from clinical childhood asthma cases and healthy controls, and determined c-CBL expression in CD4 + T cells. Asthma was induced in neonatal mice by ovalbumin (OVA) intraperitoneal injection and aerosol inhalation, and c-CBL expression in CD4 + T cells from peripheral blood and spleen was measured. Gain-of-function experiments was performed to confirm the effects of c-CBL on Th2 differentiation in vitro. Finally, c-CBL was delivered into asthmatic mice via lentivirus infection to verify its effects on experimental asthma.
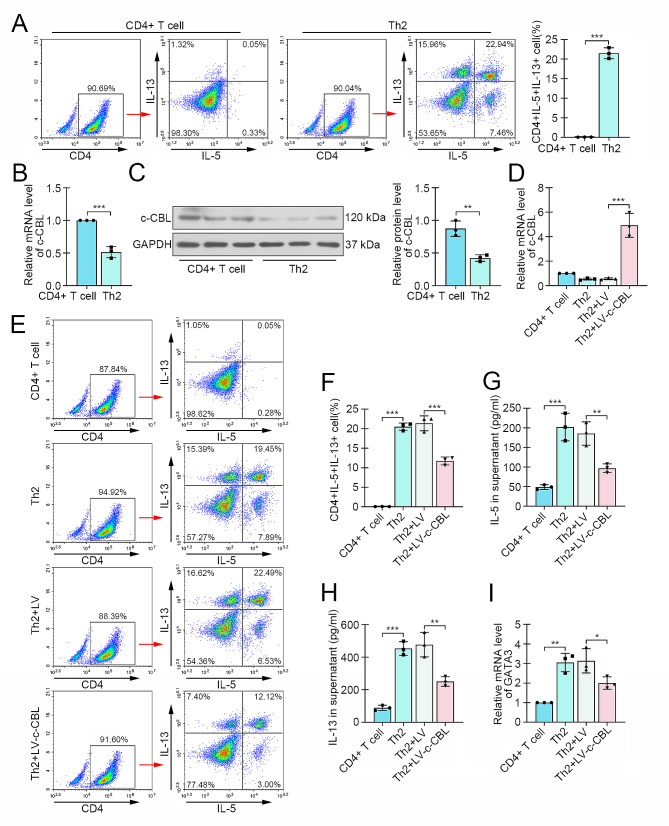
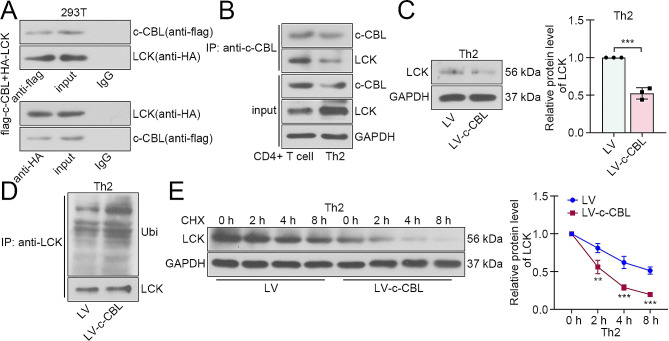
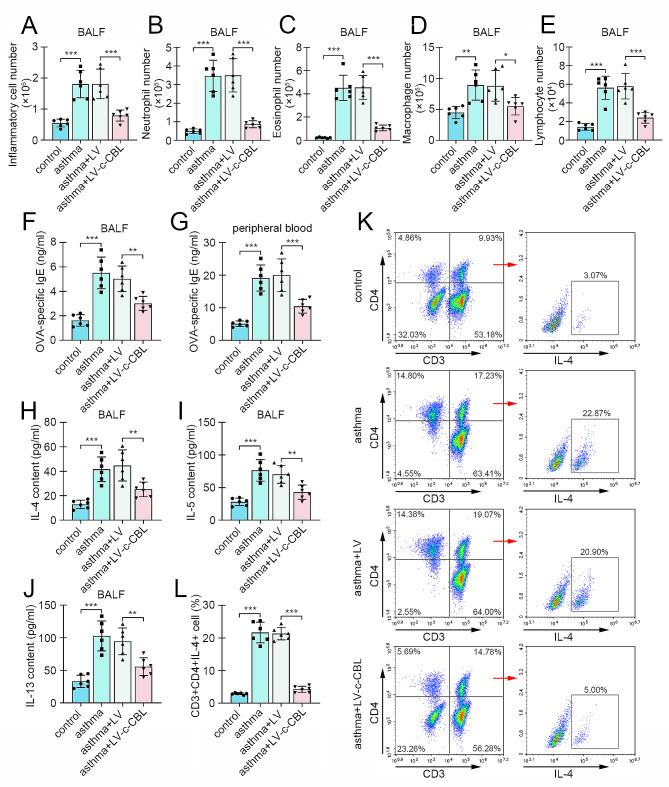
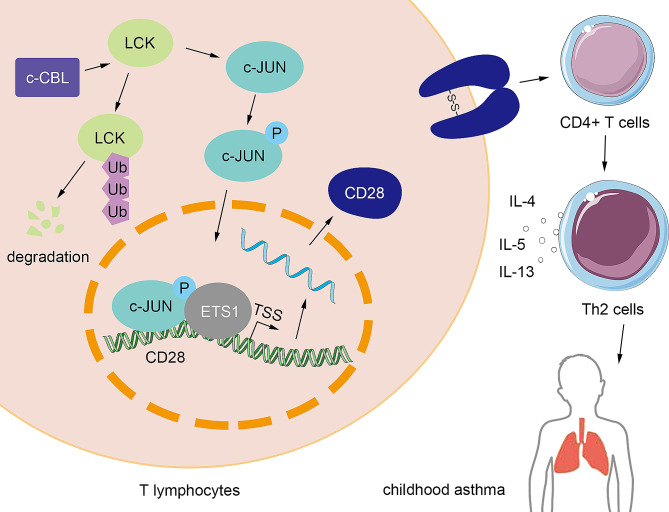
Results: c-CBL was lowly expressed in CD4 + T cells from asthmatic children than those of healthy controls. Similarly, it was downregulated in CD4 + T cells from peripheral blood and spleen of asthma mice. Overexpression of c-CBL restrained lung pathological injury and type 2 inflammation in experimental asthmatic mice. Gain-of-function experiments demonstrated that c-CBL inhibited Th2 differentiation of CD4 + T cells from healthy children, and mediated the ubiquitination of lymphocyte cell-specific protein-tyrosine kinase (LCK). LCK acted as a kinase to phosphorylate and activate c-JUN, which was predicted to bind promoter sequence of CD28 by bioinformatic analysis. Dual-luciferase reporter assay verified that c-JUN and ETS1 synergically enhanced transcription of CD28, and this transcription activation was aggravated by LCK overexpression.
Conclusion: c-CBL alleviated asthma and suppressed Th2 differentiation by facilitating LCK ubiquitination, interrupting c-JUN activation and CD28 expression in vivo and in vitro. c-CBL/LCK/c-JUN/ETS1/CD28 axis was partially involved in childhood asthma, and may provide novel insights for clinical treatment for asthma.
Keywords: Asthma; Inflammation; LCK; Th2 cells; c-CBL.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Downregulation of miRNA-451a Promotes the Differentiation of CD4+ T Cells towards Th2 Cells by Upregulating ETS1 in Childhood Asthma.J Innate Immun. 2021;13(1):38-48. doi: 10.1159/000509714. Epub 2020 Dec 3. J Innate Immun. 2021. PMID: 33271553 Free PMC article.
-
E3 ubiquitin ligase Cbl-b suppresses proallergic T cell development and allergic airway inflammation.Cell Rep. 2014 Feb 27;6(4):709-23. doi: 10.1016/j.celrep.2014.01.012. Epub 2014 Feb 6. Cell Rep. 2014. PMID: 24508458 Free PMC article.
-
Silencing TRIM8 alleviates allergic asthma and suppressing Th2 differentiation through inhibiting NF-κB/NLRP3 signaling pathway.Immunol Lett. 2024 Dec;270:106923. doi: 10.1016/j.imlet.2024.106923. Epub 2024 Sep 10. Immunol Lett. 2024. PMID: 39260527
-
Critical Involvement of CD44 in T Helper Type 2 Cell-Mediated Eosinophilic Airway Inflammation in a Mouse Model of Acute Asthma.Front Immunol. 2022 Jan 7;12:811600. doi: 10.3389/fimmu.2021.811600. eCollection 2021. Front Immunol. 2022. PMID: 35069598 Free PMC article. Review.
-
The IFITM protein family in adaptive immunity.Immunology. 2020 Apr;159(4):365-372. doi: 10.1111/imm.13163. Epub 2019 Dec 22. Immunology. 2020. PMID: 31792954 Free PMC article. Review.
Cited by
-
Bioinformatics analysis and preliminary validation of autophagy-related genes in asthma disease.Sci Rep. 2025 Jul 1;15(1):21475. doi: 10.1038/s41598-025-08316-4. Sci Rep. 2025. PMID: 40596380 Free PMC article.
-
The Ubiquitin-Proteasome System in Asthma: Mechanisms and Therapeutic Possibilities.Clin Rev Allergy Immunol. 2025 Sep 11;68(1):86. doi: 10.1007/s12016-025-09081-y. Clin Rev Allergy Immunol. 2025. PMID: 40935955 Review.
References
-
- Alqarni SA, Bineid A, Ahmad SF, Al-Harbi NO, Alqahtani F, Ibrahim KE, et al. Blockade of tyrosine kinase, LCK leads to reduction in Airway Inflammation through Regulation of Pulmonary Th2/Treg Balance and oxidative stress in Cockroach Extract-Induced mouse model of allergic asthma. Metabolites. 2022;12(9). 10.3390/metabo12090793. indexed in Pubmed: 36144198. - PMC - PubMed
-
- Alqarni SA, Ahmad SF, Alqahtani F, Al-Harbi NO, Alshehri S, Ibrahim KE, et al. Inhibition of non-receptor tyrosine kinase LCK partially mitigates mixed granulocytic airway inflammation in a murine model of asthma. Int Immunopharmacol. 2023;119(110225). 10.1016/j.intimp.2023.110225. indexed in Pubmed: 37119678. - PubMed
-
- Bacharier LB, Maspero JF, Katelaris CH, Fiocchi AG, Gagnon R, de Mir I, et al. Dupilumab in children with uncontrolled moderate-to-severe asthma. N Engl J Med. 2021;385(24):2230–40. doi: 10.1056/NEJMoa2106567, indexed in Pubmed: 34879449. - PubMed
-
- Bassuk AG, Leiden JM. A direct physical association between ETS and AP-1 transcription factors in normal human T cells. Immunity. 1995;3(2):223–37. 10.1016/1074-7613. )90092-6, indexed in Pubmed: 7648395. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
