

AIBP controls TLR4 inflammarafts and mitochondrial dysfunction in a mouse model of Alzheimer's disease
- PMID: 39342323
- PMCID: PMC11439205
- DOI: 10.1186/s12974-024-03214-4
AIBP controls TLR4 inflammarafts and mitochondrial dysfunction in a mouse model of Alzheimer's disease
Abstract
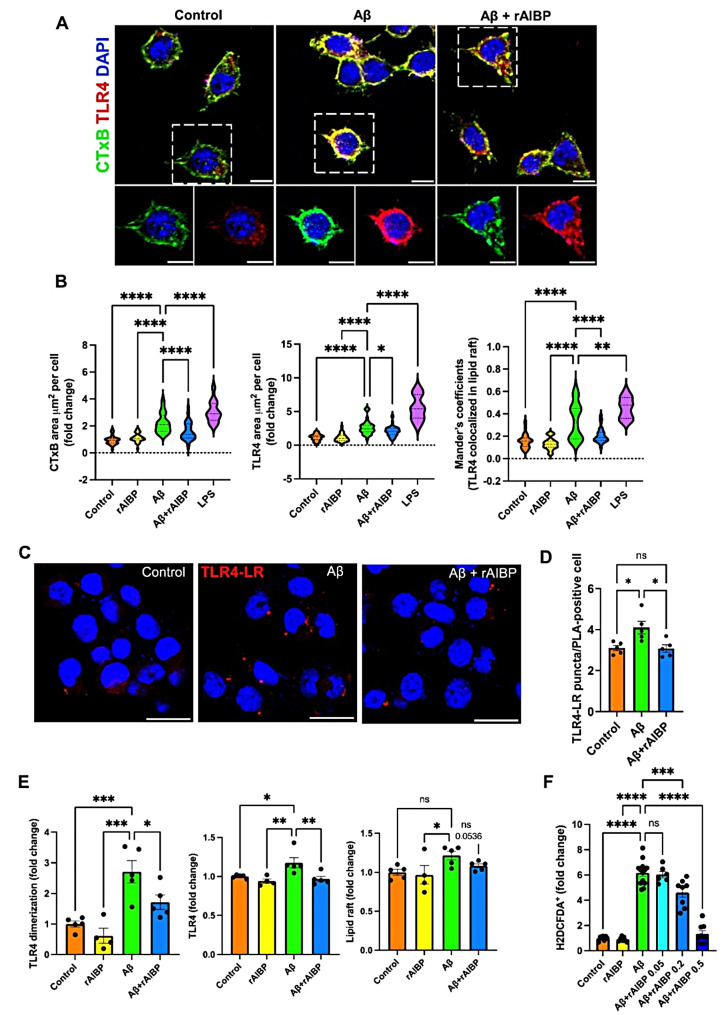
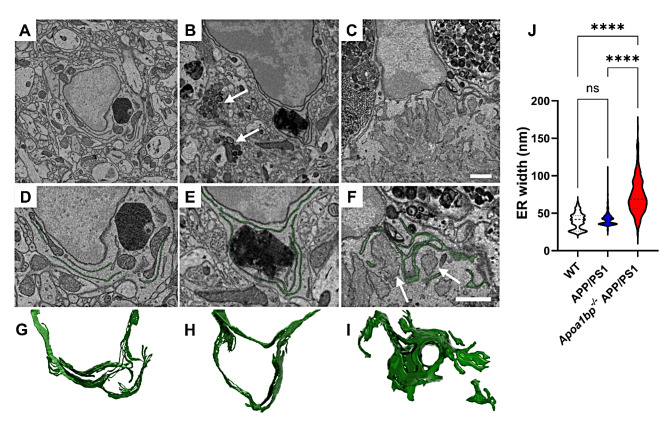
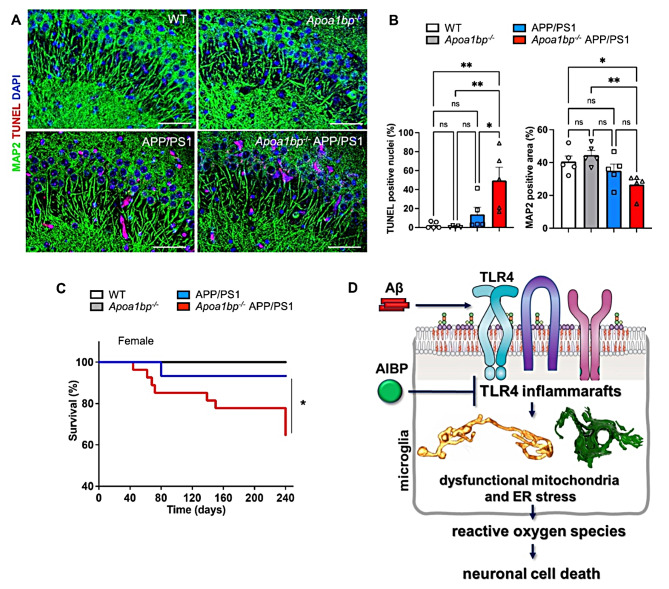
Microglia-driven neuroinflammation plays an important role in the development of Alzheimer's disease. Microglia activation is accompanied by the formation and chronic expression of TLR4 inflammarafts, defined as enlarged and cholesterol-rich lipid rafts serving as an assembly platform for TLR4 dimers and complexes of other inflammatory receptors. The secreted apoA-I binding protein (APOA1BP or AIBP) binds TLR4 and selectively targets cholesterol depletion machinery to TLR4 inflammaraft-expressing inflammatory, but not homeostatic microglia. Here we demonstrated that amyloid-beta (Aβ) induced formation of TLR4 inflammarafts in microglia in vitro and in the brain of APP/PS1 mice. Mitochondria in Apoa1bp-/- APP/PS1 microglia were hyperbranched and cupped, which was accompanied by increased reactive oxygen species and the dilated endoplasmic reticulum. The size and number of Aβ plaques and neuronal cell death were significantly increased, and the animal survival was decreased in Apoa1bp-/-APP/PS1 compared to APP/PS1 female mice. These results suggest that AIBP exerts control of TLR4 inflammarafts and mitochondrial dynamics in microglia and plays a protective role in Alzheimer's disease associated oxidative stress and neurodegeneration.
Keywords: AIBP; Alzheimer’s disease; Inflammaraft; Lipid rafts; Microglia; Mitochondria; Neuroinflammation; Oxidative stress; TLR4.
© 2024. The Author(s).
Conflict of interest statement
Y.I.M., S.-H.C. and W.-K.J. are inventors listed in patent applications related to the topic of this paper. Y.I.M. is scientific co-founder of Raft Pharmaceuticals LLC. The terms of this arrangement have been reviewed and approved by the University of California, San Diego in accordance with its conflict of interest policies. Other authors declare that they have no competing interests.
Figures





Update of
-
AIBP controls TLR4 inflammarafts and mitochondrial dysfunction in a mouse model of Alzheimer's disease.bioRxiv [Preprint]. 2024 Mar 27:2024.02.16.580751. doi: 10.1101/2024.02.16.580751. bioRxiv. 2024. Update in: J Neuroinflammation. 2024 Sep 28;21(1):245. doi: 10.1186/s12974-024-03214-4. PMID: 38586011 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
