

Over-activation of iNKT cells aggravate lung injury in bronchopulmonary dysplasia mice
- PMID: 39342744
- PMCID: PMC11470607
- DOI: 10.1016/j.redox.2024.103370
Over-activation of iNKT cells aggravate lung injury in bronchopulmonary dysplasia mice
Abstract
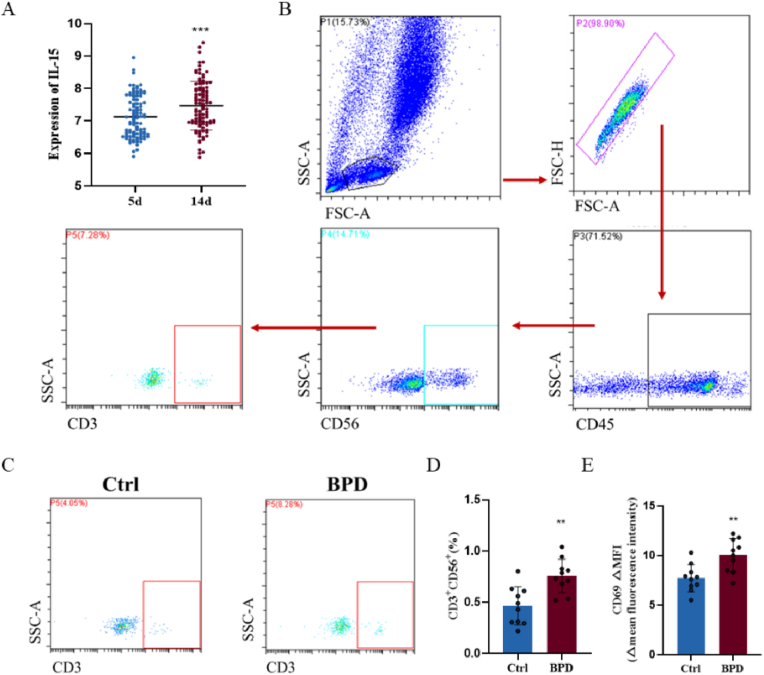
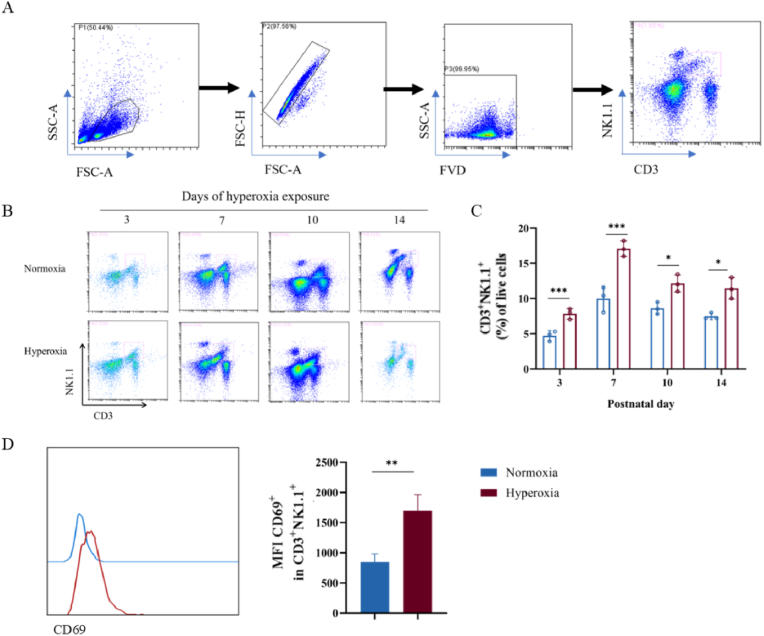
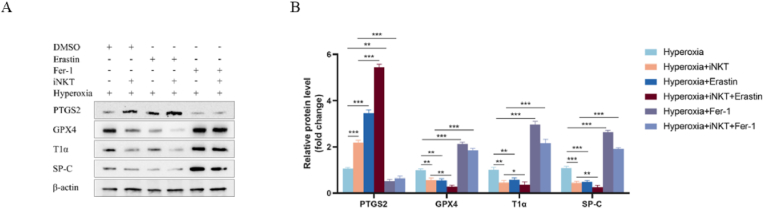
Bronchopulmonary dysplasia (BPD) is a severe lung disease in preterm infants, the abnormal proliferate and differentiate ability of type II epithelial cells (AEC II) is the key to the pathological basis of BPD. Mechanisms regarding abnormal AEC II in BPD remain unclear. The present work investigated the role and mechanisms of invariant natural killer T (iNKT) cells in lung disorder in BPD using public datasets, clinical samples, a hyperoxia-induced BPD mouse model and AEC II-iNKT cells transwell co-culture system. Firstly, we found that the NKT cells development factor IL-15 increased over time in patients with BPD in public databases, and clinically collected peripheral blood NKT cells in patients with BPD were increased. Subsequently, the percentage of iNKT cells increased in hyperoxia group compared with normoxia group, with the highest at P7, accompanied by increased activation with abnormal lung development. The administration of anti-CD1d neutralizing antibody to inhibit iNKT cells could alleviate the abnormal lung development of hyperoxia group mice, while α-GalCer administration could aggravate lung injury in hyperoxia group mice, and adoptive transfer of iNKT cells could aggravate the abnormal lung development in hyperoxia group mice. In addition, to further verify the role of iNKT cells on AEC II, AEC II-iNKT cells co-culture system was established. The presence of iNKT cells could aggravate the abnormal expression of SP-C and T1α under hyperoxia. Meanwhile, RNA-seq analysis showed that ferroptosis-related genes were highly expressed in AEC II co-cultured with iNKT cells under hyperoxia. We further validated the effect of the presence of iNKT cells under hyperoxia environment on AEC II ferroptosis levels, suggested that iNKT cells promote AEC II ferroptosis under hyperoxia, accompanied by decreased expression of SP-C and T1α. Our study found that the recruitment of iNKT cells in the lung may be an important cause of alveolarization disorder in BPD.
Keywords: Alveolar type II epithelial cells; Bronchopulmonary dysplasia; Differentiation; Ferroptosis; Invariant natural killer T cells.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures






References
-
- Bancalari E., Jain D. Bronchopulmonary dysplasia: 50 Years after the original description. Neonatology. 2019;115(4):384–391. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
