

A missense mutation in the barley Xan-h gene encoding the Mg-chelatase subunit I leads to a viable pale green line with reduced daily transpiration rate
- PMID: 39343835
- PMCID: PMC11439855
- DOI: 10.1007/s00299-024-03328-2
A missense mutation in the barley Xan-h gene encoding the Mg-chelatase subunit I leads to a viable pale green line with reduced daily transpiration rate
Abstract
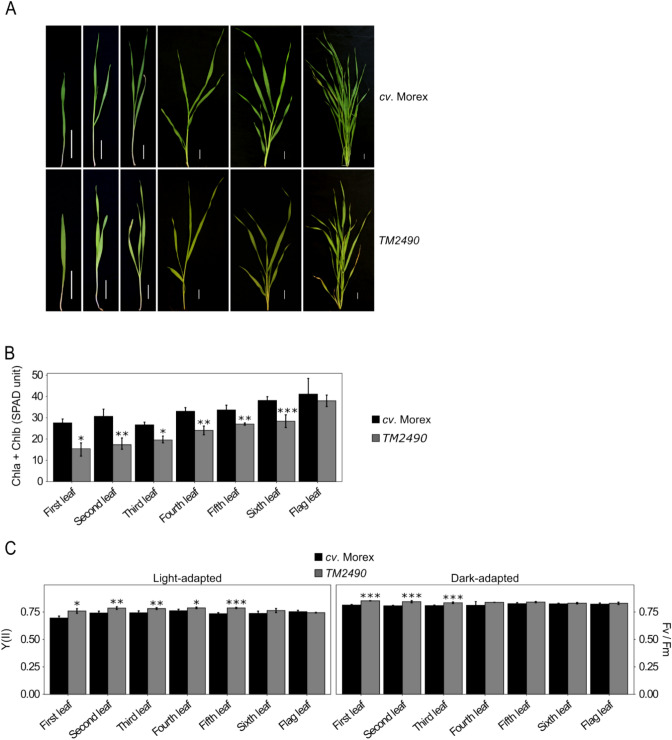
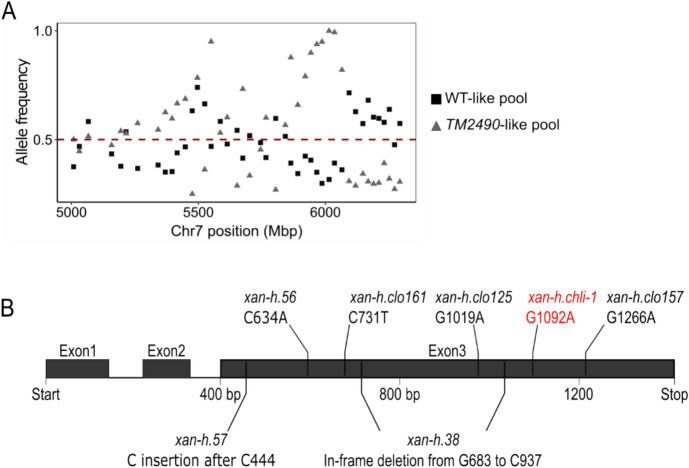
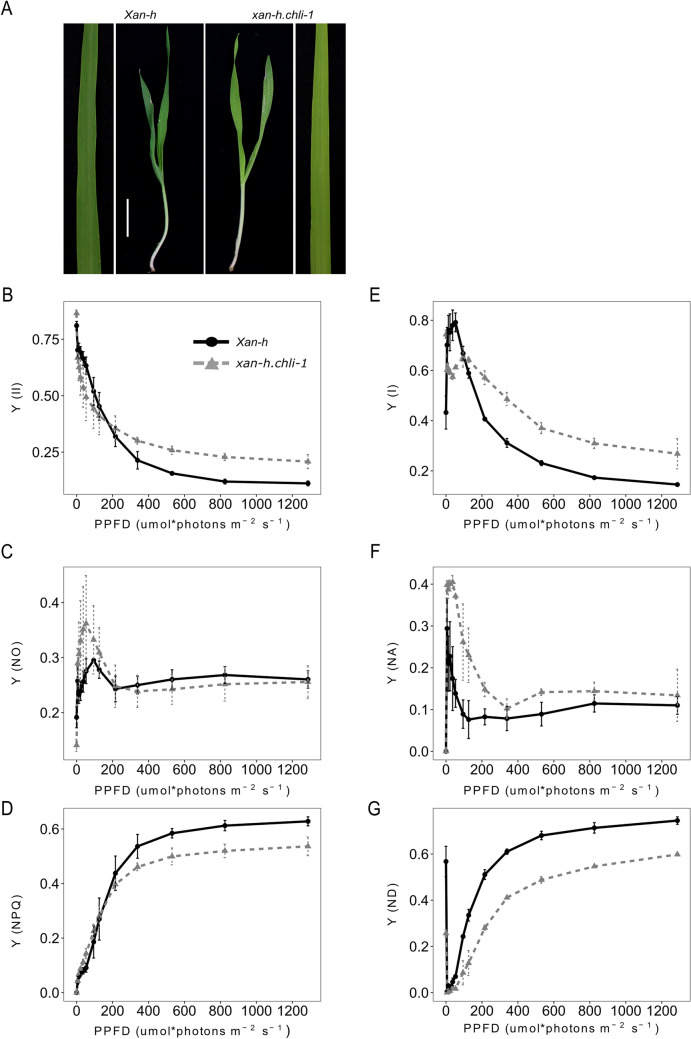
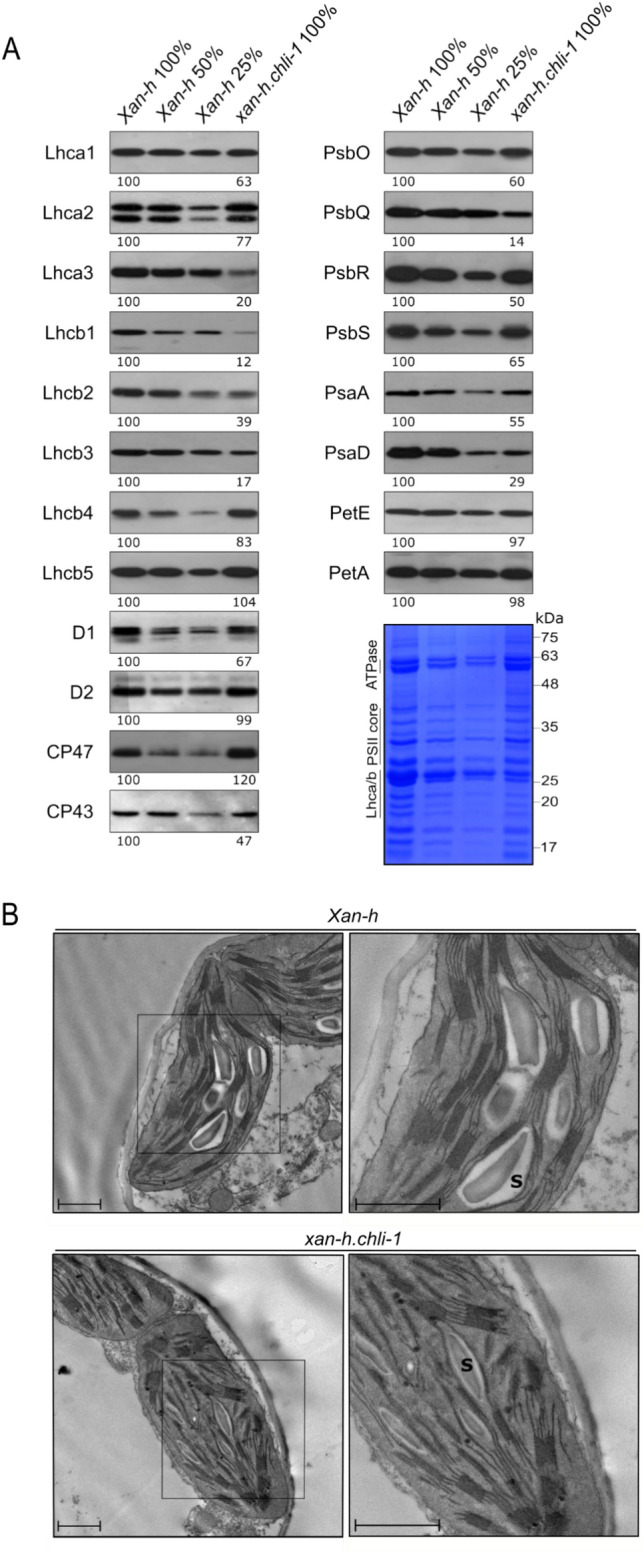
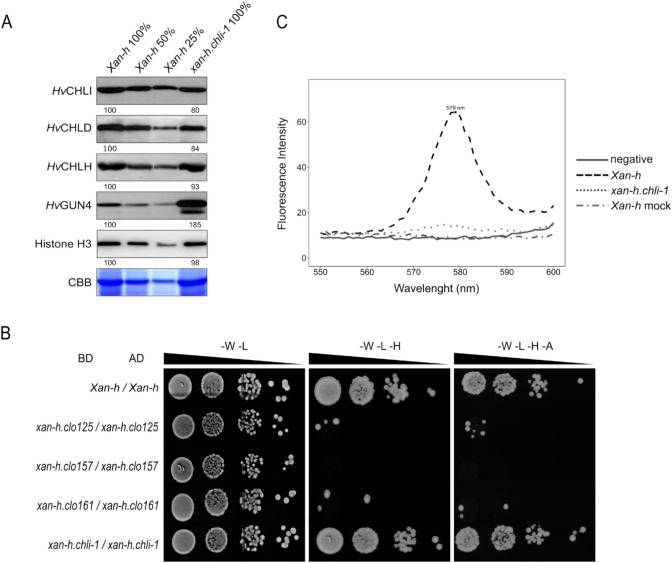
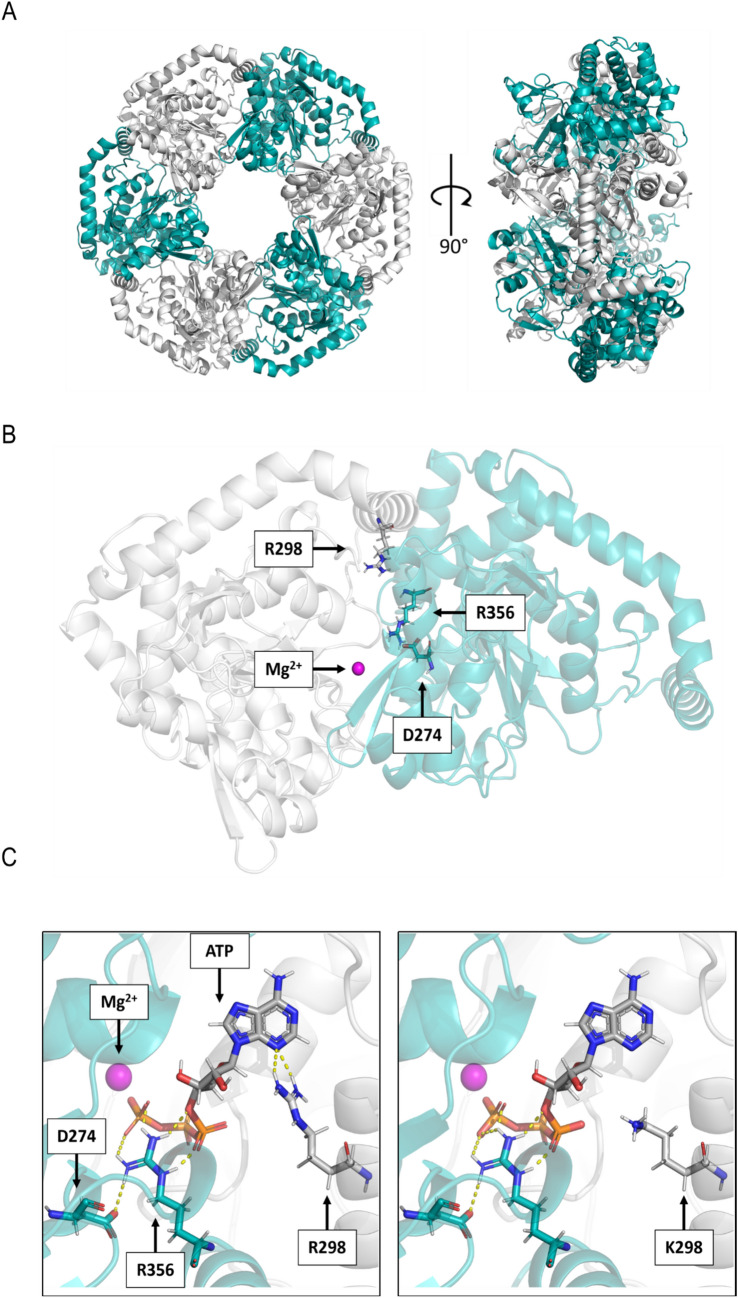
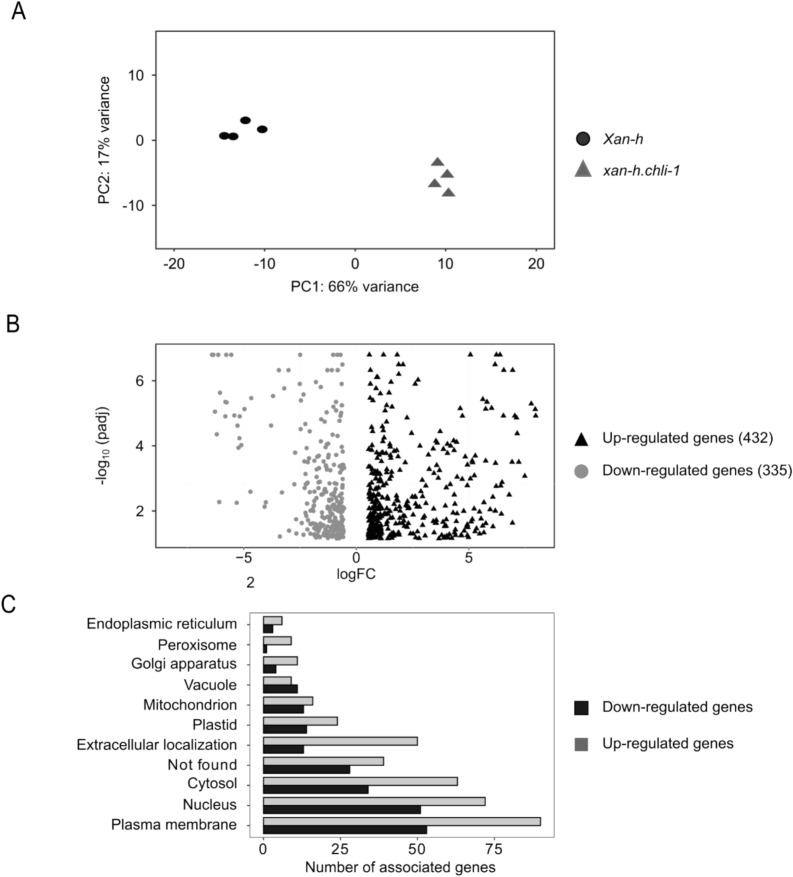
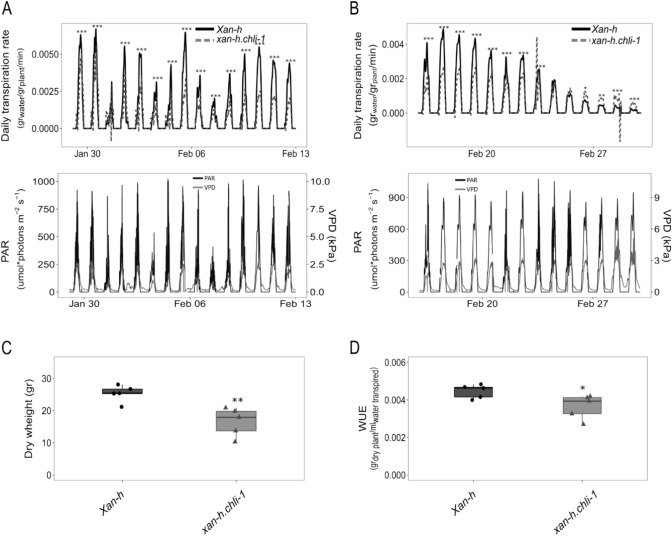
The barley mutant xan-h.chli-1 shows phenotypic features, such as reduced leaf chlorophyll content and daily transpiration rate, typical of wild barley accessions and landraces adapted to arid climatic conditions. The pale green trait, i.e. reduced chlorophyll content, has been shown to increase the efficiency of photosynthesis and biomass accumulation when photosynthetic microorganisms and tobacco plants are cultivated at high densities. Here, we assess the effects of reducing leaf chlorophyll content in barley by altering the chlorophyll biosynthesis pathway (CBP). To this end, we have isolated and characterised the pale green barley mutant xan-h.chli-1, which carries a missense mutation in the Xan-h gene for subunit I of Mg-chelatase (HvCHLI), the first enzyme in the CBP. Intriguingly, xan-h.chli-1 is the only known viable homozygous mutant at the Xan-h locus in barley. The Arg298Lys amino-acid substitution in the ATP-binding cleft causes a slight decrease in HvCHLI protein abundance and a marked reduction in Mg-chelatase activity. Under controlled growth conditions, mutant plants display reduced accumulation of antenna and photosystem core subunits, together with reduced photosystem II yield relative to wild-type under moderate illumination, and consistently higher than wild-type levels at high light intensities. Moreover, the reduced content of leaf chlorophyll is associated with a stable reduction in daily transpiration rate, and slight decreases in total biomass accumulation and water-use efficiency, reminiscent of phenotypic features of wild barley accessions and landraces that thrive under arid climatic conditions.
Keywords: Barley; Canopy photosynthesis; Chlorophyll biosynthesis; Drought stress; Mg-chelatase; Pale green leaves.
© 2024. The Author(s).
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures
References
-
- Barbato R, Tadini L, Cannata R, Peracchio C, Jeran N, Alboresi A, Morosinotto T et al (2020) Higher order photoprotection mutants reveal the importance of ΔpH-dependent photosynthesis-control in preventing light induced damage to both photosystem II and photosystem I. Sci Rep 10(1):1–14. 10.1038/s41598-020-62717-1 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 1032 17/06/2022/Next-Generation EU: PIANO NAZIONALE DI RIPRESA E RESILIENZA
- CN00000022/Next-Generation EU: PIANO NAZIONALE DI RIPRESA E RESILIENZA
- Italy-Israel Joint Call for Proposals on Scientific/Ministero degli Affari Esteri e della Cooperazione Internazionale
- Technological Cooperation 2021/Ministero degli Affari Esteri e della Cooperazione Internazionale
- 101082091 - BEST-CROP/HORIZON EUROPE Food, Bioeconomy, Natural Resources, Agriculture and Environment
