

Developmental maturation and regional heterogeneity but no sexual dimorphism of the murine CNS myelin proteome
- PMID: 39344832
- PMCID: PMC11660532
- DOI: 10.1002/glia.24614
Developmental maturation and regional heterogeneity but no sexual dimorphism of the murine CNS myelin proteome
Abstract
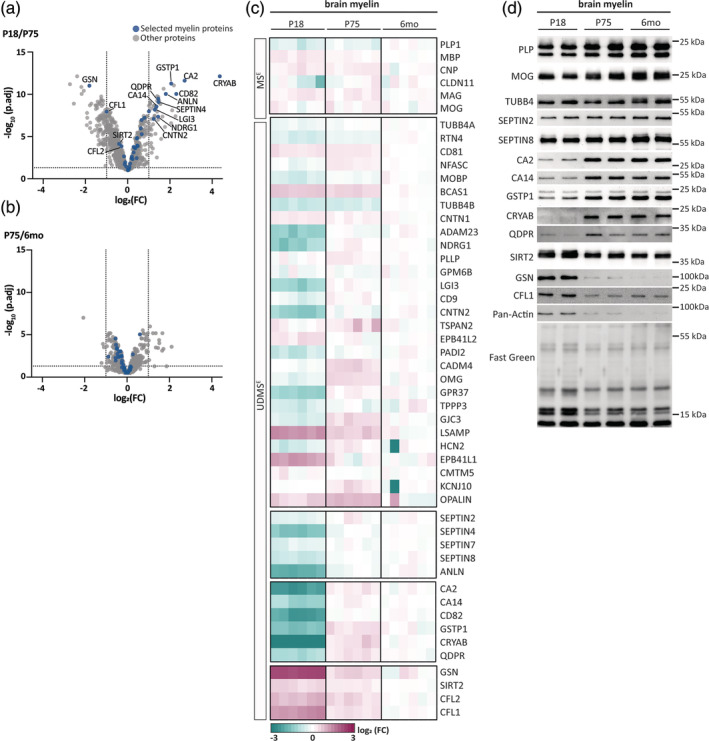
The molecules that constitute myelin are critical for the integrity of axon/myelin-units and thus speed and precision of impulse propagation. In the CNS, the protein composition of oligodendrocyte-derived myelin has evolutionarily diverged and differs from that in the PNS. Here, we hypothesized that the CNS myelin proteome also displays variations within the same species. We thus used quantitative mass spectrometry to compare myelin purified from mouse brains at three developmental timepoints, from brains of male and female mice, and from four CNS regions. We find that most structural myelin proteins are of approximately similar abundance across all tested conditions. However, the abundance of multiple other proteins differs markedly over time, implying that the myelin proteome matures between P18 and P75 and then remains relatively constant until at least 6 months of age. Myelin maturation involves a decrease of cytoskeleton-associated proteins involved in sheath growth and wrapping, along with an increase of all subunits of the septin filament that stabilizes mature myelin, and of multiple other proteins which potentially exert protective functions. Among the latter, quinoid dihydropteridine reductase (QDPR) emerges as a highly specific marker for mature oligodendrocytes and myelin. Conversely, female and male mice display essentially similar myelin proteomes. Across the four CNS regions analyzed, we note that spinal cord myelin exhibits a comparatively high abundance of HCN2-channels, required for particularly long sheaths. These findings show that CNS myelination involves developmental maturation of myelin protein composition, and regional differences, but absence of evidence for sexual dimorphism.
Keywords: axon/glia‐interaction; myelin maturation; oligodendrocyte; quantitative proteomics; white matter.
© 2024 The Author(s). GLIA published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Conservation and divergence of myelin proteome and oligodendrocyte transcriptome profiles between humans and mice.Elife. 2022 May 11;11:e77019. doi: 10.7554/eLife.77019. Elife. 2022. PMID: 35543322 Free PMC article.
-
N-Wasp Regulates Oligodendrocyte Myelination.J Neurosci. 2020 Aug 5;40(32):6103-6111. doi: 10.1523/JNEUROSCI.0912-20.2020. Epub 2020 Jun 29. J Neurosci. 2020. PMID: 32601246 Free PMC article.
-
Mammalian target of rapamycin promotes oligodendrocyte differentiation, initiation and extent of CNS myelination.J Neurosci. 2014 Mar 26;34(13):4453-65. doi: 10.1523/JNEUROSCI.4311-13.2014. J Neurosci. 2014. PMID: 24671992 Free PMC article.
-
Myelin sheaths: glycoproteins involved in their formation, maintenance and degeneration.Cell Mol Life Sci. 2002 Nov;59(11):1851-71. doi: 10.1007/pl00012510. Cell Mol Life Sci. 2002. PMID: 12530518 Free PMC article. Review.
-
The oligodendrocyte growth cone and its actin cytoskeleton: A fundamental element for progenitor cell migration and CNS myelination.Glia. 2020 Jul;68(7):1329-1346. doi: 10.1002/glia.23735. Epub 2019 Nov 7. Glia. 2020. PMID: 31696982 Free PMC article. Review.
Cited by
-
Myelin Lipid Composition in the Central Nervous System Is Regionally Distinct and Requires Mechanistic Target of Rapamycin Signaling.Glia. 2025 Sep;73(9):1841-1859. doi: 10.1002/glia.70042. Epub 2025 May 26. Glia. 2025. PMID: 40417825 Free PMC article.
-
Prominosomes - a particular class of extracellular vesicles containing prominin-1/CD133?J Nanobiotechnology. 2025 Jan 29;23(1):61. doi: 10.1186/s12951-025-03102-w. J Nanobiotechnology. 2025. PMID: 39881297 Free PMC article. Review.
References
-
- Alvarez‐Sanchez, N. , & Dunn, S. E. (2023). Immune cell contributors to the female sex bias in multiple sclerosis and experimental autoimmune encephalomyelitis. Current Topics in Behavioral Neurosciences, 62, 333–373. - PubMed
-
- Autilio, L. A. , Norton, W. T. , & Terry, R. D. (1964). The preparation and some properties of purified myelin from the central nervous system. Journal of Neurochemistry, 11, 17–27. - PubMed
-
- Benjamini, Y. , & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series B: Statistical Methodology, 57, 289–300.
-
- Buscham, T. J. , Eichel‐Vogel, M. A. , Steyer, A. M. , Jahn, O. , Strenzke, N. , Dardawal, R. , Memhave, T. R. , Siems, S. B. , Müller, C. , Meschkat, M. , Sun, T. , Ruhwedel, T. , Möbius, W. , Krämer‐Albers, E.‐M. , Boretius, S. , Nave, K.‐A. , & Werner, H. B. (2022). Progressive axonopathy when oligodendrocytes lack the myelin protein CMTM5. eLife, 11, e75523. - PMC - PubMed
-
- Cammer, W. , Bieler, L. , Fredman, T. , & Norton, W. T. (1977). Quantitation of myelin carbonic anhydrase‐development and subfractionation of rat brain myelin and comparison with myelin from other species. Brain Research, 138, 17–28. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
