

Tetracistronic minigenomes elucidate a functional promoter for Ghana virus and unveils Cedar virus replicase promiscuity for all henipaviruses
- PMID: 39345144
- PMCID: PMC11495047
- DOI: 10.1128/jvi.00806-24
Tetracistronic minigenomes elucidate a functional promoter for Ghana virus and unveils Cedar virus replicase promiscuity for all henipaviruses
Abstract
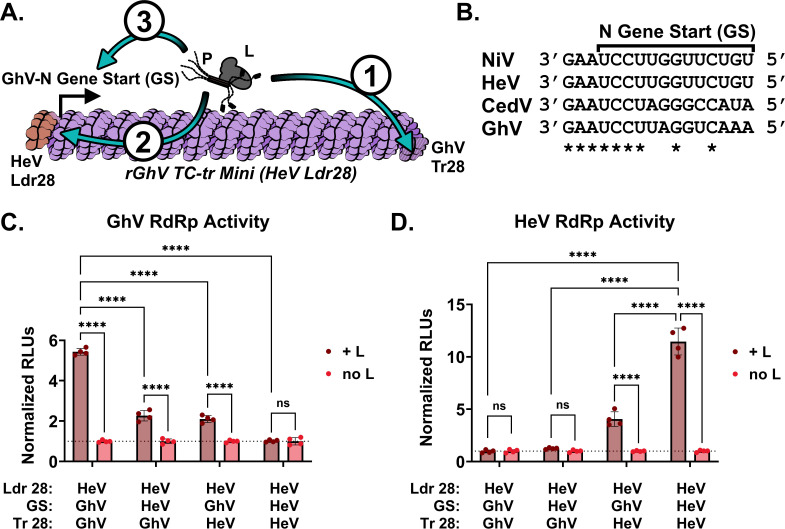
Batborne henipaviruses, such as Nipah and Hendra viruses, represent a major threat to global health due to their propensity for spillover, severe pathogenicity, and high mortality rate in human hosts. Coupled with the absence of approved vaccines or therapeutics, work with the prototypical species and uncharacterized, emergent species is restricted to high biocontainment facilities. There is a scarcity of such specialized spaces for research, and often, the scope and capacity of research, which can be conducted at BSL-4, is limited. Therefore, there is a pressing need for innovative life-cycle modeling systems to enable comprehensive research within lower biocontainment settings. This work showcases tetracistronic, transcription, and replication-competent minigenomes for the Nipah, Hendra, and Cedar viruses, which encode viral proteins facilitating budding, fusion, and receptor binding. We validate the functionality of all encoded viral proteins and demonstrate a variety of applications to interrogate the viral life cycle. Notably, we found that the Cedar virus replicase exhibits remarkable promiscuity, efficiently driving replication and transcription of minigenomes from all tested henipaviruses. We also apply this technology to Ghana virus (GhV), an emergent species that has so far not been isolated in culture. We demonstrate that the reported sequence of GhV is incomplete, but that this missing sequence can be substituted with analogous sequences from other henipaviruses. The use of our GhV system establishes the functionality of the GhV replicase and identifies two antivirals that are highly efficacious against the GhV polymerase.
Importance: Henipaviruses are recognized as significant global health threats due to their high mortality rates and lack of effective vaccines or therapeutics. Due to the requirement for high biocontainment facilities, the scope of research which may be conducted on henipaviruses is limited. To address this challenge, we developed innovative tetracistronic, transcription, and replication-competent minigenomes. We demonstrate that these systems replicate key aspects of the viral life cycle, such as budding, fusion, and receptor binding, and are safe for use in lower biocontainment settings. Importantly, the application of this system to the Ghana virus revealed that its known sequence is incomplete; however, substituting the missing sequences with those from other henipaviruses allowed us to overcome this challenge. We demonstrate that the Ghana virus replicative machinery is functional and can identify two orally efficacious antivirals effective against it. Our research offers a versatile system for life-cycle modeling of highly pathogenic henipaviruses at low biocontainment.
Keywords: Ghana virus; Nipah virus; antivirals; emerging pathogens; henipavirus; high biocontainment; minigenome; paramyxovirus; reverse genetics; viral RNA dependent RNA polymerase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Update of
-
Tetracistronic Minigenomes Elucidate a Functional Promoter for Ghana Virus and Unveils Cedar Virus Replicase Promiscuity for all Henipaviruses.bioRxiv [Preprint]. 2024 Apr 16:2024.04.16.589704. doi: 10.1101/2024.04.16.589704. bioRxiv. 2024. Update in: J Virol. 2024 Oct 22;98(10):e0080624. doi: 10.1128/jvi.00806-24. PMID: 38659760 Free PMC article. Updated. Preprint.
References
-
- Wells HL, Loh E, Nava A, Solorio MR, Lee MH, Lee J, Sukor JRA, Navarrete-Macias I, Liang E, Firth C, Epstein JH, Rostal MK, Zambrana-Torrelio C, Murray K, Daszak P, Goldstein T, Mazet JAK, Lee B, Hughes T, Durigon E, Anthony SJ. 2022. Classification of new morbillivirus and jeilongvirus sequences from bats sampled in Brazil and Malaysia. Arch Virol 167:1977–1987. doi:10.1007/s00705-022-05500-z - DOI - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
- T32 AI007647/AI/NIAID NIH HHS/United States
- AI07647/HHS | National Institutes of Health (NIH)
- AI060549/HHS | National Institutes of Health (NIH)
- AI071002/HHS | National Institutes of Health (NIH)
- AI149033/HHS | National Institutes of Health (NIH)
- R01 AI071002/AI/NIAID NIH HHS/United States
- AI171403/HHS | National Institutes of Health (NIH)
- T32 AI060549/AI/NIAID NIH HHS/United States
- 1842169/NSF | National Science Foundation Graduate Research Fellowship Program (GRFP)
- INV-048877/Bill and Melinda Gates Foundation (GF)
- U19 AI171403/AI/NIAID NIH HHS/United States
- R21 AI149033/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
