

PhoPQ-mediated lipopolysaccharide modification governs intrinsic resistance to tetracycline and glycylcycline antibiotics in Escherichia coli
- PMID: 39345149
- PMCID: PMC11495068
- DOI: 10.1128/msystems.00964-24
PhoPQ-mediated lipopolysaccharide modification governs intrinsic resistance to tetracycline and glycylcycline antibiotics in Escherichia coli
Abstract
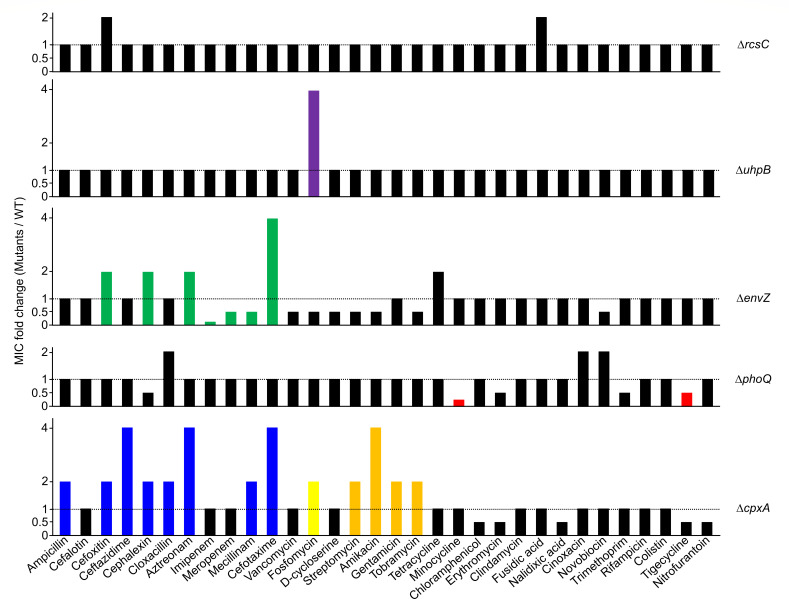
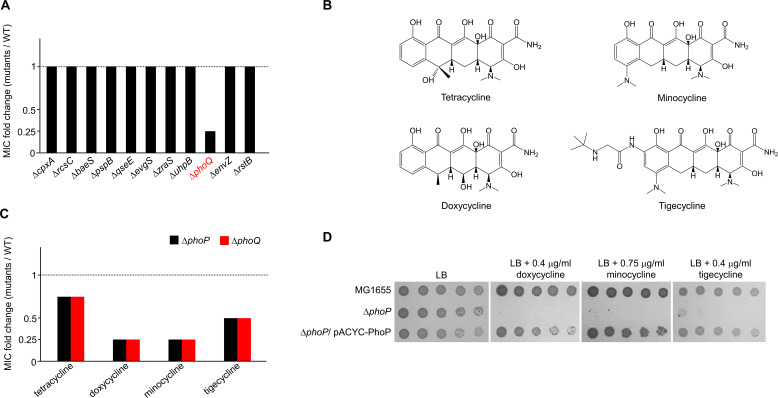
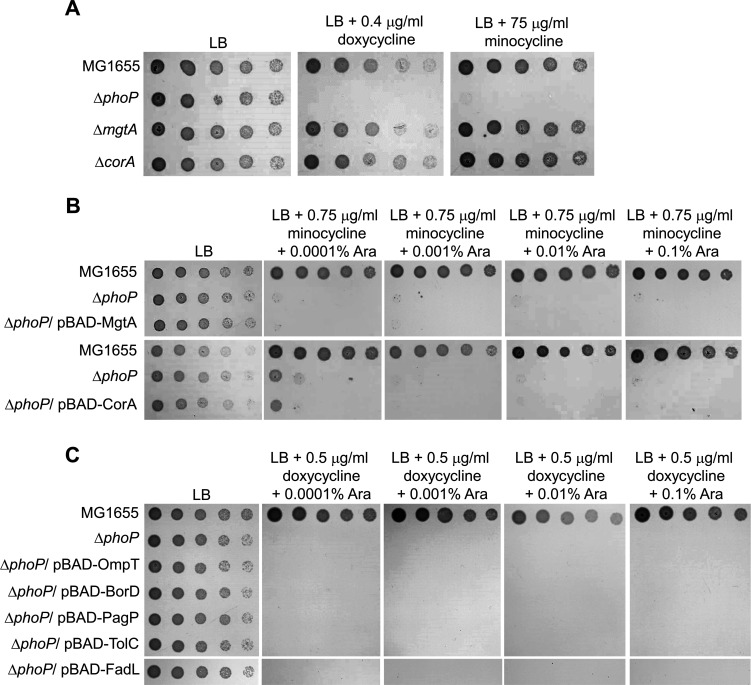
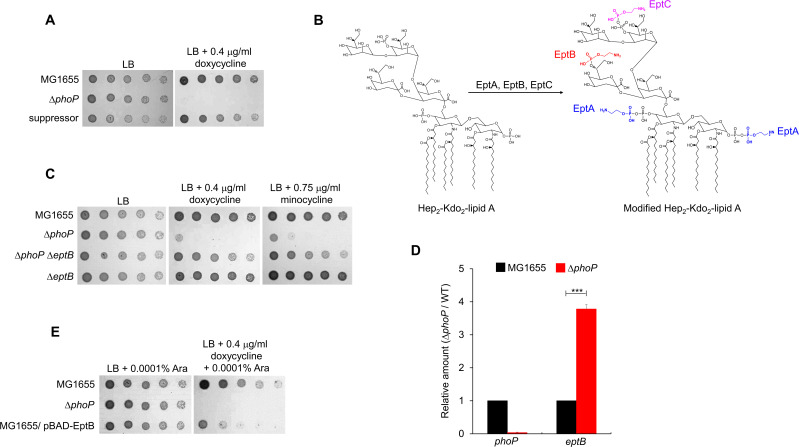
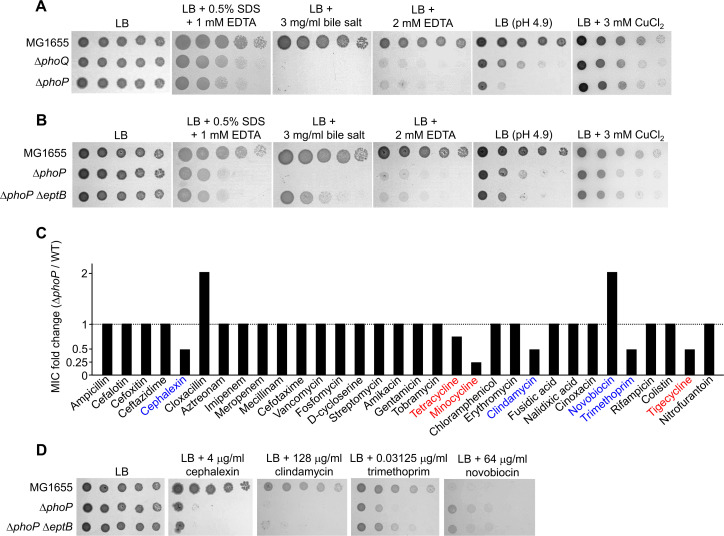
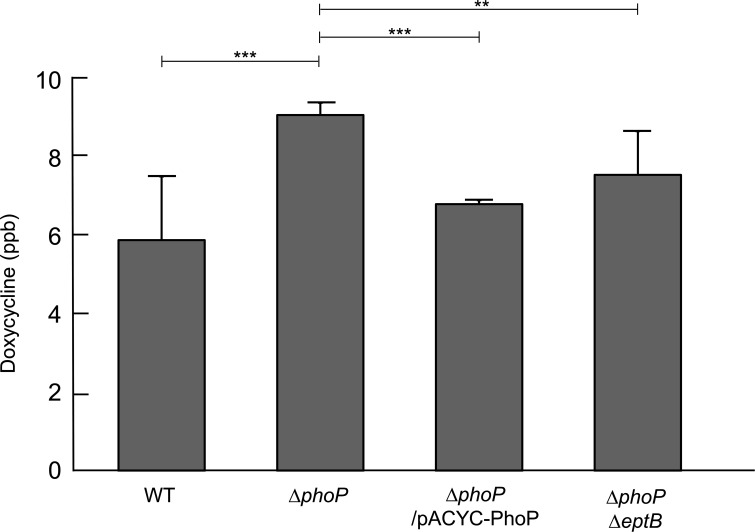
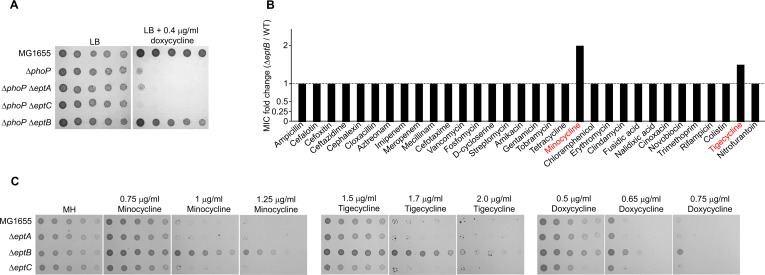
Tetracyclines and glycylcycline are among the important antibiotics used to combat infections caused by multidrug-resistant Gram-negative pathogens. Despite the clinical importance of these antibiotics, their mechanisms of resistance remain unclear. In this study, we elucidated a novel mechanism of resistance to tetracycline and glycylcycline antibiotics via lipopolysaccharide (LPS) modification. Disruption of the Escherichia coli PhoPQ two-component system, which regulates the transcription of various genes involved in magnesium transport and LPS modification, leads to increased susceptibility to tetracycline, minocycline, doxycycline, and tigecycline. These phenotypes are caused by enhanced expression of phosphoethanolamine transferase EptB, which catalyzes the modification of the inner core sugar of LPS. PhoPQ-mediated regulation of EptB expression appears to affect the intracellular transportation of doxycycline. Disruption of EptB increases resistance to tetracycline and glycylcycline antibiotics, whereas the other two phosphoethanolamine transferases, EptA and EptC, that participate in the modification of other LPS residues, are not associated with resistance to tetracyclines and glycylcycline. Overall, our results demonstrated that PhoPQ-mediated modification of a specific residue of LPS by phosphoethanolamine transferase EptB governs intrinsic resistance to tetracycline and glycylcycline antibiotics.
Importance: Elucidating the resistance mechanisms of clinically important antibiotics helps in maintaining the clinical efficacy of antibiotics and in the prescription of adequate antibiotic therapy. Although tetracycline and glycylcycline antibiotics are clinically important in combating multidrug-resistant Gram-negative bacterial infections, their mechanisms of resistance are not fully understood. Our research demonstrates that the E. coli PhoPQ two-component system affects resistance to tetracycline and glycylcycline antibiotics by controlling the expression of phosphoethanolamine transferase EptB, which catalyzes the modification of the inner core residue of lipopolysaccharide (LPS). Therefore, our findings highlight a novel resistance mechanism to tetracycline and glycylcycline antibiotics and the physiological significance of LPS core modification in E. coli.
Keywords: PhoPQ; antibiotic resistance; doxycycline; glycylcycline; lipopolysaccharide modification; minocycline; tetracycline; tigecycline.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Subinhibitory concentration stress of colistin enhanced PhoPQ expression in Escherichia coli harboring mcr-1.J Basic Microbiol. 2021 Nov;61(11):1029-1034. doi: 10.1002/jobm.202100355. Epub 2021 Sep 12. J Basic Microbiol. 2021. PMID: 34510495
-
In vitro and in vivo antibacterial activities of a novel glycylcycline, the 9-t-butylglycylamido derivative of minocycline (GAR-936).Antimicrob Agents Chemother. 1999 Apr;43(4):738-44. doi: 10.1128/AAC.43.4.738. Antimicrob Agents Chemother. 1999. PMID: 10103174 Free PMC article.
-
Diacylglycerol Kinase A Is Essential for Polymyxin Resistance Provided by EptA, MCR-1, and Other Lipid A Phosphoethanolamine Transferases.J Bacteriol. 2022 Feb 15;204(2):e0049821. doi: 10.1128/JB.00498-21. Epub 2021 Nov 29. J Bacteriol. 2022. PMID: 34843376 Free PMC article.
-
Tetracycline antibiotics and resistance mechanisms.Biol Chem. 2014 May;395(5):559-75. doi: 10.1515/hsz-2013-0292. Biol Chem. 2014. PMID: 24497223 Review.
-
Part X. Tetracycline and glycylcycline antimicrobials.J Okla State Med Assoc. 2007 Jun;100(6):241-7. J Okla State Med Assoc. 2007. PMID: 17708423 Review. No abstract available.
Cited by
-
A historical perspective on the multifunctional outer membrane channel protein TolC in Escherichia coli.NPJ Antimicrob Resist. 2025 Jan 25;3(1):6. doi: 10.1038/s44259-025-00078-3. NPJ Antimicrob Resist. 2025. PMID: 39863731 Free PMC article. Review.
-
Breaking Barriers: Exploiting Envelope Biogenesis and Stress Responses to Develop Novel Antimicrobial Strategies in Gram-Negative Bacteria.Pathogens. 2024 Oct 11;13(10):889. doi: 10.3390/pathogens13100889. Pathogens. 2024. PMID: 39452760 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
