

Chlamydia-driven ISG15 expression dampens the immune response of epithelial cells independently of ISGylation
- PMID: 39345209
- PMCID: PMC11559041
- DOI: 10.1128/mbio.02401-24
Chlamydia-driven ISG15 expression dampens the immune response of epithelial cells independently of ISGylation
Abstract
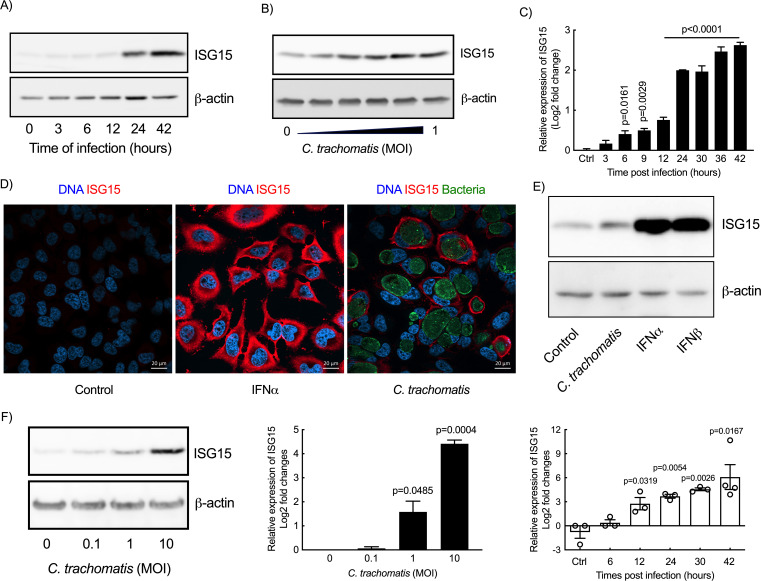
Excessive inflammation upon Chlamydia trachomatis infection can cause severe damages in the female genital tract. This obligate intracellular bacterium develops mainly in epithelial cells, whose innate response contributes to the overall inflammatory response to infection. The ubiquitin-like protein interferon-stimulated gene 15 (ISG15) stimulates interferon γ (IFNγ) production and is required for bacterial clearance in several infectious contexts. Here, we describe and investigate the consequences of the increase in ISG15 expression by epithelial cells infected with C. trachomatis. Infection of HeLa cells and primary ecto-cervical epithelial cells resulted in a transcriptional upregulation of ISG15 expression. This did not involve the canonical type I interferon (IFN-I) signaling pathway and depended instead on the activation of the STING/TBK1/IRF3 pathway. The absence or reduction of ISG15 synthesis led to increased production of several cytokines and chemokines, including interleukin (IL) 6 and IL8. This implicates that ISG15 normally dampens the immune response induced by C. trachomatis infection in epithelial cells. ISG15 exerted its control from an intracellular location, but without involving ISGylation. Finally, higher levels of inflammation and delayed bacterial clearance were observed in the genital tracts of ISG15-KO mice infected by C. trachomatis compared with wild-type animals; however, IFNγ production was unchanged. Altogether, our data show that ISG15 expression acts as a brake on the immune response to C. trachomatis infection in epithelial cells and limits bacterial burden and inflammation in mice.IMPORTANCEInfection of epithelial cells by Chlamydia trachomatis elicits an innate immune response by these cells. The signaling pathways involved, and their outcomes, are still very poorly understood. In this paper, we described how Chlamydia infection triggered the expression of ISG15, a small molecule normally associated to type I interferon (IFN-I) signaling and control of INF-γ production. ISG15 synthesis by epithelial cells attenuated their immune response to Chlamydia infection. In mice, we observed that ISG15 displayed a marginal role in modulating the production of IFN-γ, a key component of the host immune response to infection, but facilitated bacterial clearance. Overall, our study strengthens the importance of ISG15 not only in the resolution of viral but also of bacterial infection and document its role of "immune brake" in the context of Chlamydia infection.
Keywords: C. trachomatis; ISGylation; epithelial cells; inflammation; innate defense; interferon stimulated gene 15 (ISG15).
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Secreted ISG15 induced by Chlamydia trachomatis infection exerts immunomodulatory effects on IFN-γ defense and inflammation.PLoS Pathog. 2025 Jul 8;21(7):e1013315. doi: 10.1371/journal.ppat.1013315. eCollection 2025 Jul. PLoS Pathog. 2025. PMID: 40627782 Free PMC article.
-
Chlamydia trachomatis modulates the expression of JAK-STAT signaling components to attenuate the type II interferon response of epithelial cells.mBio. 2024 Oct 16;15(10):e0183424. doi: 10.1128/mbio.01834-24. Epub 2024 Aug 28. mBio. 2024. PMID: 39194253 Free PMC article.
-
Chlamydia trachomatis Is Resistant to Inclusion Ubiquitination and Associated Host Defense in Gamma Interferon-Primed Human Epithelial Cells.mBio. 2016 Dec 13;7(6):e01417-16. doi: 10.1128/mBio.01417-16. mBio. 2016. PMID: 27965446 Free PMC article.
-
Restriction and evasion: a review of IFNγ-mediated cell-autonomous defense pathways during genital Chlamydia infection.Pathog Dis. 2024 Feb 7;82:ftae019. doi: 10.1093/femspd/ftae019. Pathog Dis. 2024. PMID: 39210512 Free PMC article. Review.
-
Male genital tract immune response against Chlamydia trachomatis infection.Reproduction. 2017 Oct;154(4):R99-R110. doi: 10.1530/REP-16-0561. Reproduction. 2017. PMID: 28878094 Review.
Cited by
-
Secreted ISG15 induced by Chlamydia trachomatis infection exerts immunomodulatory effects on IFN-γ defense and inflammation.PLoS Pathog. 2025 Jul 8;21(7):e1013315. doi: 10.1371/journal.ppat.1013315. eCollection 2025 Jul. PLoS Pathog. 2025. PMID: 40627782 Free PMC article.
References
-
- Knight E, Fahey D, Cordova B, Hillman M, Kutny R, Reich N, Blomstrom D. 1988. A 15-kDa interferon-induced protein is derived by COOH-terminal processing of A 17-kDa precursor. J Biol Chem 263:4520–4522. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
