

Thalattosauria in time and space: a review of thalattosaur spatiotemporal occurrences, presumed evolutionary relationships and current ecological hypotheses
- PMID: 39345254
- PMCID: PMC11427521
- DOI: 10.1186/s13358-024-00333-6
Thalattosauria in time and space: a review of thalattosaur spatiotemporal occurrences, presumed evolutionary relationships and current ecological hypotheses
Abstract
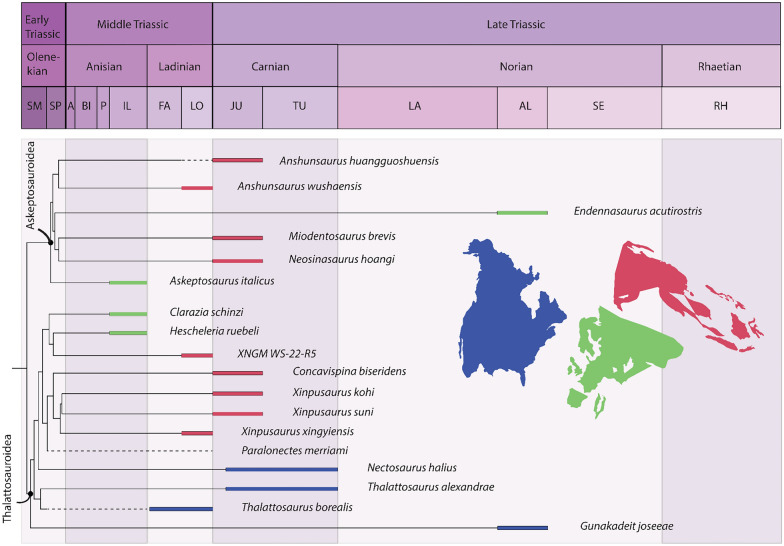
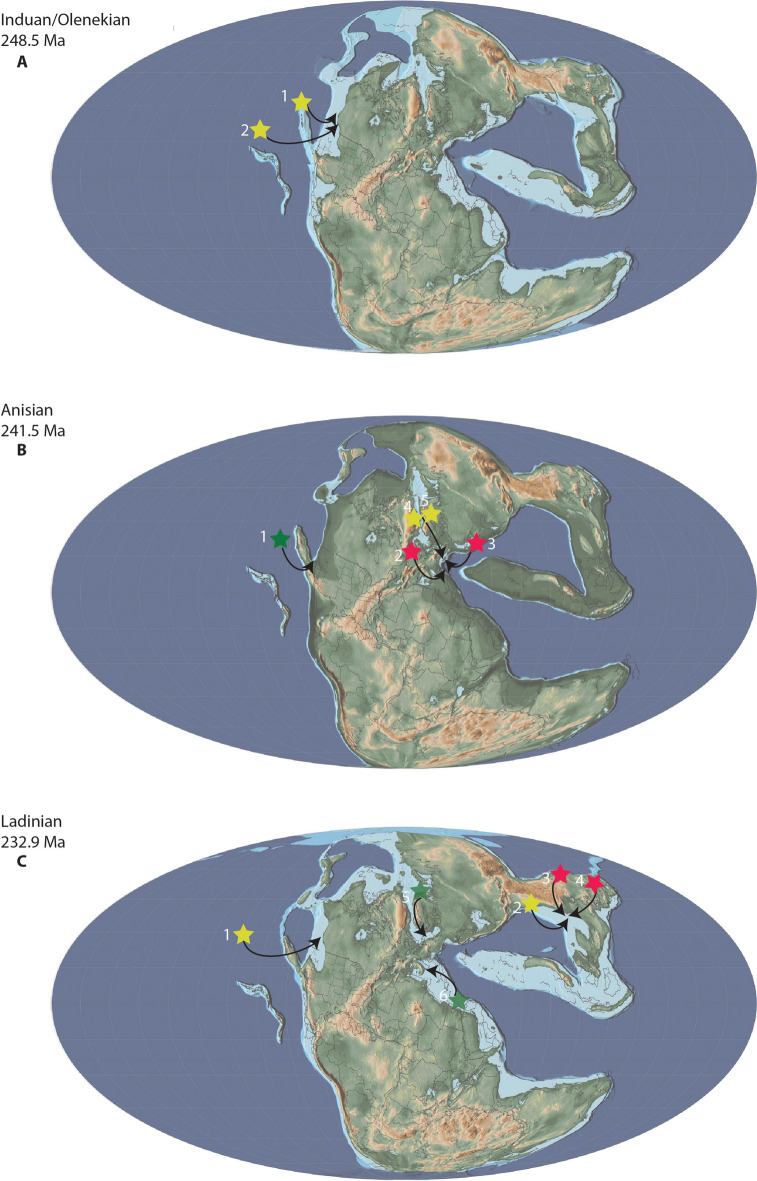
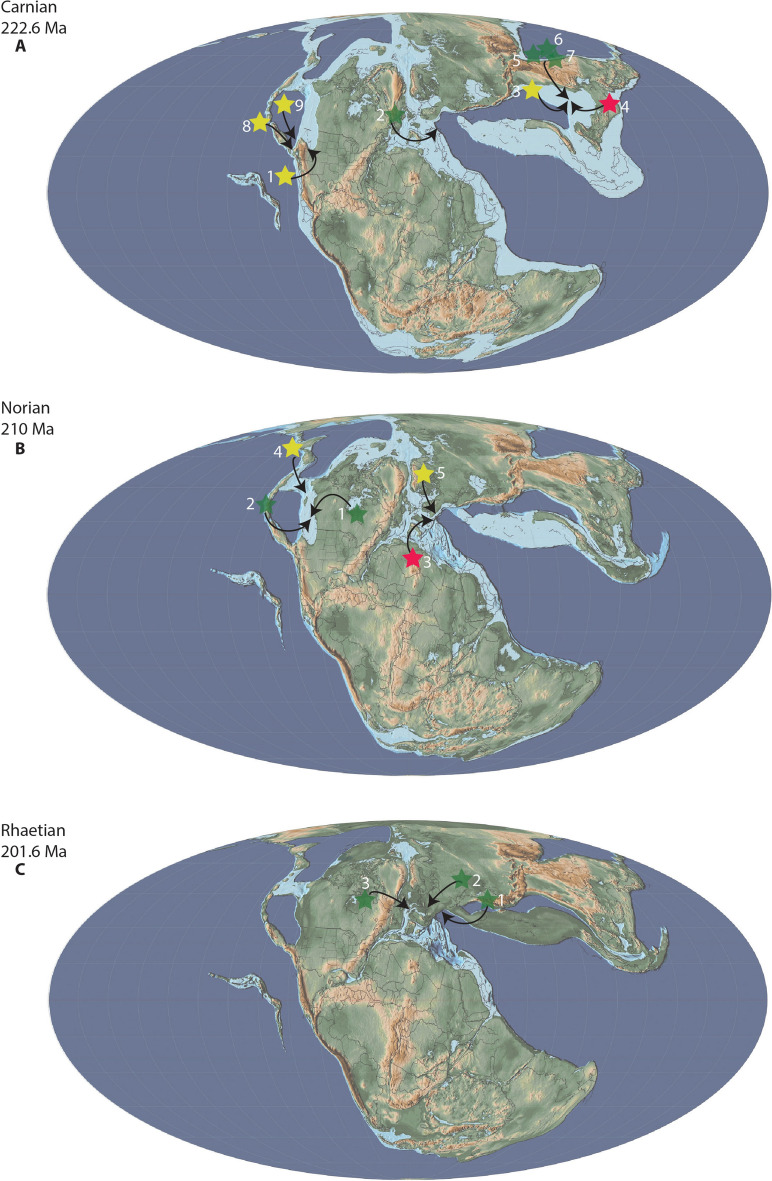
In the wake of the greatest mass extinction in Earth's history, the End-Permian Mass Extinction, the Triassic was a time of recovery and innovation. Aided by warm climatic conditions and favorable ecological circumstances, many reptilian clades originated and rapidly diversified during this time. This set the stage for numerous independent invasions of the marine realm by several reptilian clades, such as Ichthyosauriformes and Sauropterygia, shaping the oceanic ecosystems for the entire Mesozoic. Although comparatively less speciose, and temporally and latitudinally more restricted, another marine reptile clade, the Thalattosauriformes, stands out because of their unusual and highly disparate cranial, dental and skeletal morphology. Research on Thalattosauriformes has been hampered by a historic dearth of material, with the exception of rare material from Lagerstätten and highly fossiliferous localities, such as that from the UNESCO world heritage site of Monte San Giorgio. Consequently, their evolutionary origins and paleobiology remain poorly understood. The recent influx of new material from southwestern China and North America has renewed interest in this enigmatic group prompting the need for a detailed review of historic work and current views. The earliest representatives of the group may have been present from the late Early Triassic onwards in British Columbia. By the Ladinian the group had achieved a wide distribution across the northern hemisphere, spanning the eastern Panthalassic as well as the eastern and western Tethyan provinces. Distinct morphological and likely ecological differences exist between the two major clades of Thalattosauriformes, the Askeptosauroidea and the Thalattosauroidea, with the latter showing a higher degree of cranial and skeletal morphological disparity. In-group relationships remain poorly resolved beyond this bipartition. Overall, thalattosaurs may be closely related to other marine reptile groups such as ichthyopterygians and sauropterygians. However, their exact position within Diapsida remains elusive. Future focal points should utilize modern digital paleontological approaches to explore the many fragmentary specimens of otherwise poorly sampled localities.
Keywords: Biogeography; Ecology; Marine reptiles; Monte San Giorgio; Phylogeny; Thalattosauriformes; Triassic.
© The Author(s) 2024.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures




Similar articles
-
Special Issue: 100 years of scientific excavations at UNESCO World Heritage Site Monte San Giorgio and global research on Triassic marine Lagerstätten.Swiss J Palaeontol. 2024;143(1):37. doi: 10.1186/s13358-024-00328-3. Epub 2024 Oct 1. Swiss J Palaeontol. 2024. PMID: 39376472 Free PMC article.
-
An armoured marine reptile from the Early Triassic of South China and its phylogenetic and evolutionary implications.Elife. 2023 Aug 8;12:e83163. doi: 10.7554/eLife.83163. Elife. 2023. PMID: 37551884 Free PMC article.
-
The oldest record of Saurosphargiformes (Diapsida) from South China could fill an ecological gap in the Early Triassic biotic recovery.PeerJ. 2022 Jul 14;10:e13569. doi: 10.7717/peerj.13569. eCollection 2022. PeerJ. 2022. PMID: 35855428 Free PMC article.
-
Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution.Biol Rev Camb Philos Soc. 2016 Feb;91(1):106-47. doi: 10.1111/brv.12161. Epub 2014 Nov 27. Biol Rev Camb Philos Soc. 2016. PMID: 25431138 Review.
-
Rapid turnover of top predators in African terrestrial faunas around the Permian-Triassic mass extinction.Curr Biol. 2023 Jun 5;33(11):2283-2290.e3. doi: 10.1016/j.cub.2023.04.007. Epub 2023 May 22. Curr Biol. 2023. PMID: 37220743 Review.
Cited by
-
Special Issue: 100 years of scientific excavations at UNESCO World Heritage Site Monte San Giorgio and global research on Triassic marine Lagerstätten.Swiss J Palaeontol. 2024;143(1):37. doi: 10.1186/s13358-024-00328-3. Epub 2024 Oct 1. Swiss J Palaeontol. 2024. PMID: 39376472 Free PMC article.
References
-
- Adams, T. L. (2009). Deposition and taphonomy of the Hound Island Late Triassic vertebrate fauna: Fossil preservation within subaqueous gravity flows. Palaios,24, 603–615.
-
- Allemand, R., Moon, B. C., & Voeten, D. F. (2022). The paleoneurology of ichthyopterygia and sauropterygia: Diverse endocranial anatomies of secondarily aquatic diapsids. In M. T. Dozo, A. Paulina-Carabajal, T. E. Macrini, & S. Walsh (Eds.), Paleoneurology of amniotes: New directions in the study of fossil endocasts (pp. 29–77). Springer International Publishing.
-
- Bardet, N. (1994). Extinction events among Mesozoic marine reptiles. Historical Biology,7(4), 313–324.
-
- Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda-Suberbiola, X., Pérez-Garcia, A., Rage, J.-C., & Vincent, P. (2014). Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research,26(3–4), 869–887.
Publication types
LinkOut - more resources
Full Text Sources