

This is a preprint.
Preventing Inappropriate Signals Pre- and Post-Ligand Perception by a Toggle-Switch Mechanism of ERECTA
- PMID: 39345552
- PMCID: PMC11429954
- DOI: 10.1101/2024.09.20.612365
Preventing Inappropriate Signals Pre- and Post-Ligand Perception by a Toggle-Switch Mechanism of ERECTA
Update in
-
Preventing inappropriate signals pre- and post-ligand perception by a toggle switch mechanism of ERECTA.Proc Natl Acad Sci U S A. 2025 Jan 28;122(4):e2420196122. doi: 10.1073/pnas.2420196122. Epub 2025 Jan 22. Proc Natl Acad Sci U S A. 2025. PMID: 39841143 Free PMC article.
Abstract
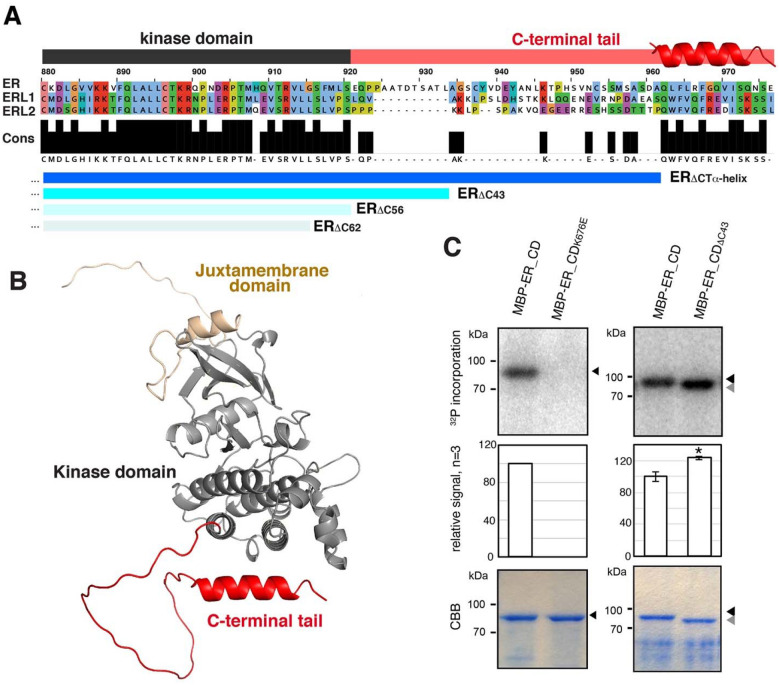
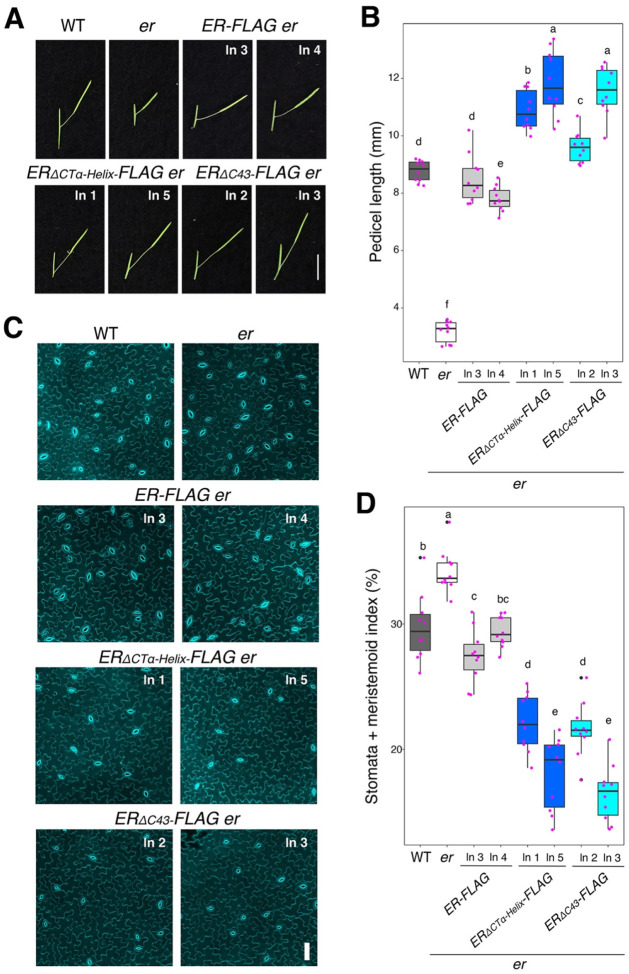
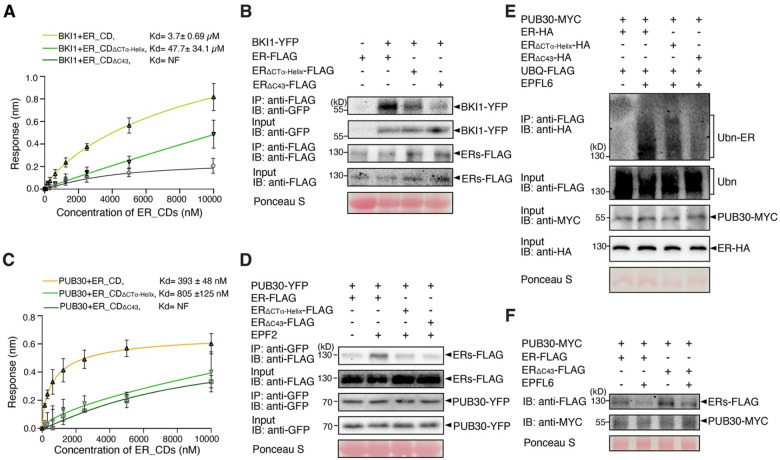
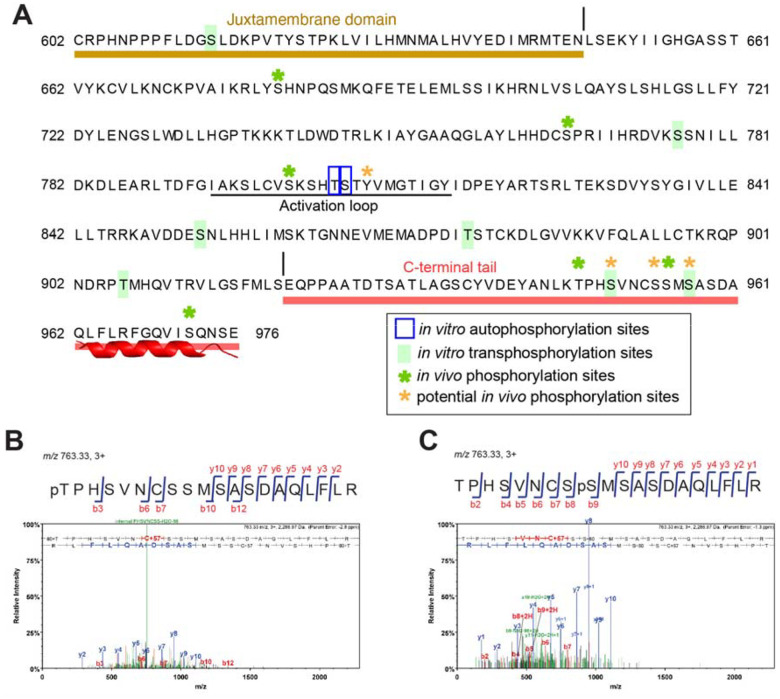
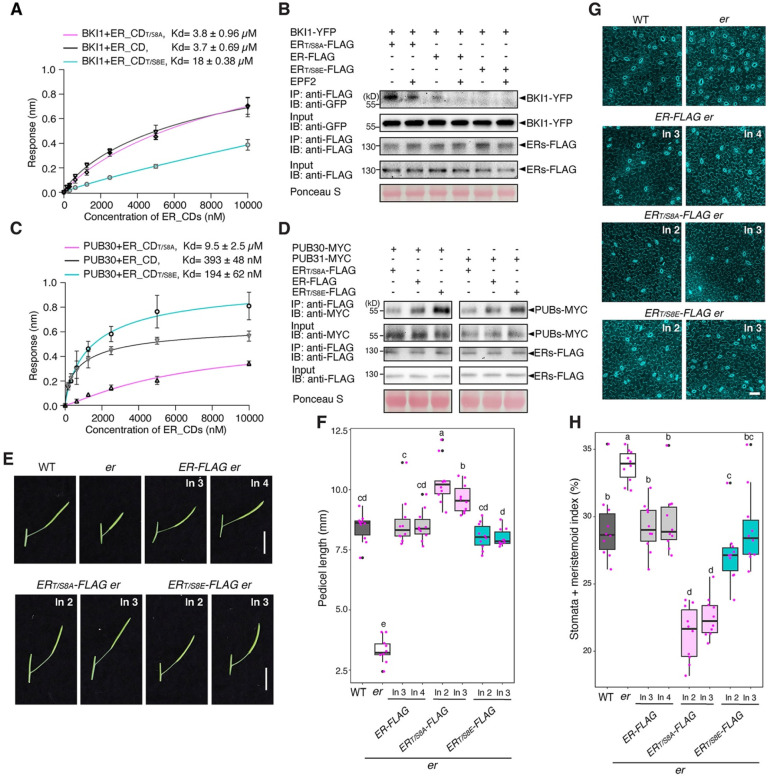
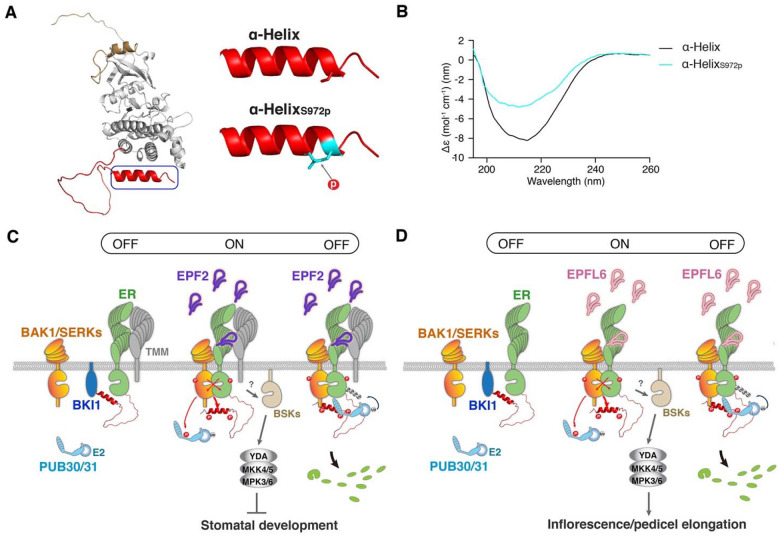
Dynamic control of signaling events requires swift regulation of receptors at an active state. By focusing on Arabidopsis ERECTA (ER) receptor kinase, which perceives peptide ligands to control multiple developmental processes, we report a mechanism preventing inappropriate receptor activity. The ER C-terminal tail (ER_CT) functions as an autoinhibitory domain: its removal confers higher kinase activity and hyperactivity during inflorescence and stomatal development. ER_CT is required for the binding of a receptor kinase inhibitor, BKI1, and two U-box E3 ligases PUB30 and PUB31 that inactivate activated ER. We further identify ER_CT as a phosphodomain transphosphorylated by the co-receptor BAK1. The phosphorylation impacts the tail structure, likely releasing from autoinhibition. The phosphonull version enhances BKI1 association, whereas the phosphomimetic version promotes PUB30/31 association. Thus, ER_CT acts as an off-on-off toggle switch, facilitating the release of BKI1 inhibition, enabling signal activation, and swiftly turning over the receptors afterwards. Our results elucidate a mechanism fine-tuning receptor signaling via a phosphoswitch module, keeping the receptor at a low basal state and ensuring the robust yet transient activation upon ligand perception.
Keywords: Arabidopsis; C-terminal Tail Domain; Phosphoregulation; Receptor Kinase; stomatal development.
Figures
References
-
- Shiu S.-H., Bleecker A. B., Plant Receptor-Like Kinase Gene Family: Diversity, Function, and Signaling. Science’s STKE 2001, re22–re22 (2001). - PubMed
-
- Morris E. R., Walker J. C., Receptor-like protein kinases: the keys to response. Current Opinion in Plant Biology 6, 339–342 (2003). - PubMed
-
- De Smet I., Voß U., Jürgens G., Beeckman T., Receptor-like kinases shape the plant. Nature Cell Biology 11, 1166 (2009). - PubMed
-
- Couto D., Zipfel C., Regulation of pattern recognition receptor signalling in plants. Nature Reviews Immunology 16, 537 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources