

This is a preprint.
A functional screen for ubiquitin regulation identifies an E3 ligase secreted by Pseudomonas aeruginosa
- PMID: 39345563
- PMCID: PMC11430079
- DOI: 10.1101/2024.09.18.613774
A functional screen for ubiquitin regulation identifies an E3 ligase secreted by Pseudomonas aeruginosa
Abstract
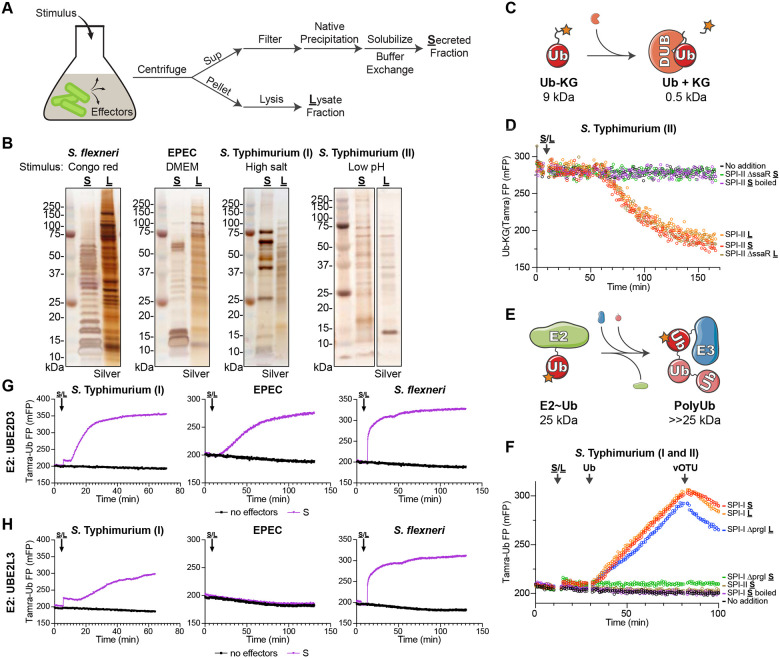
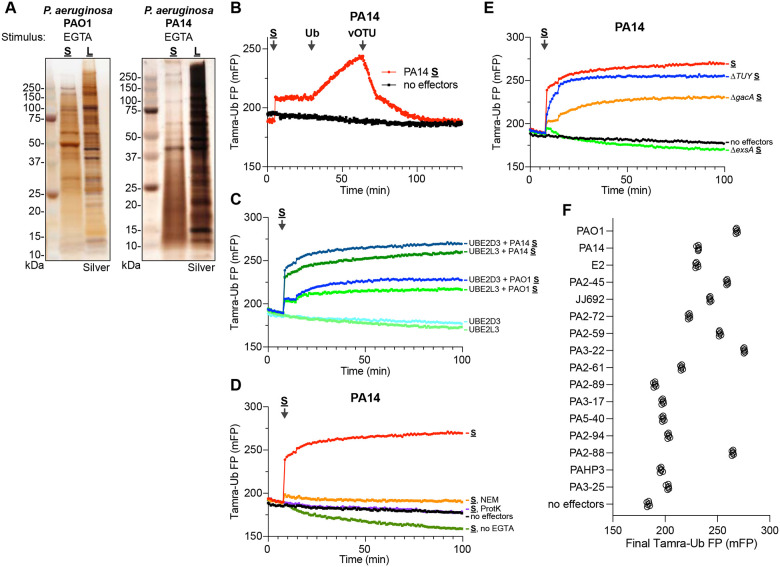
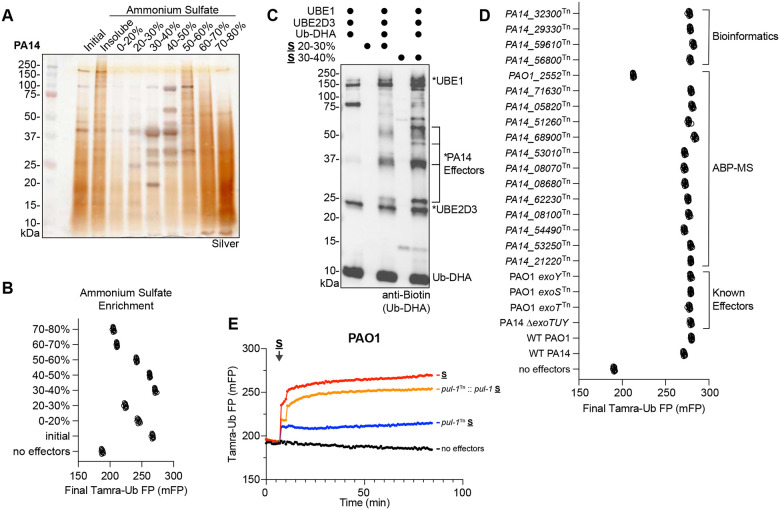
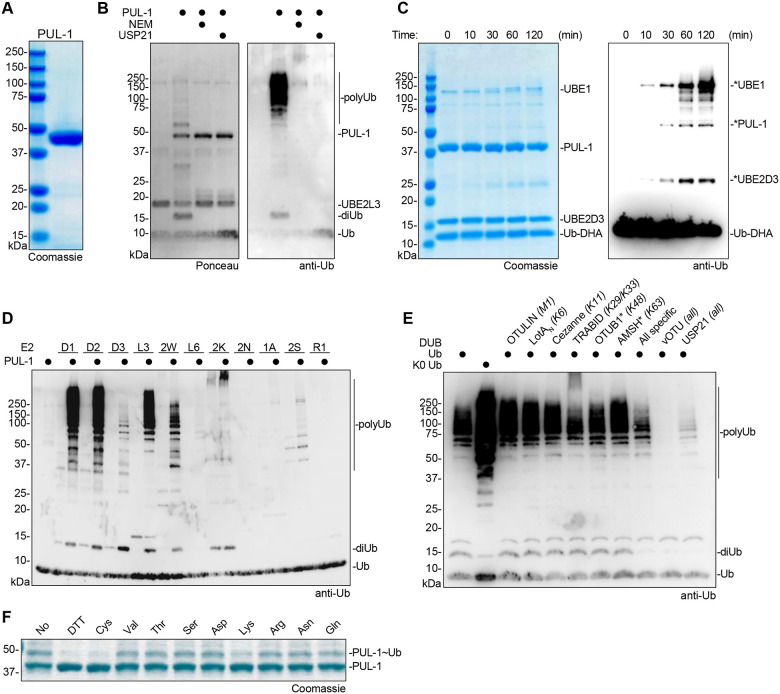
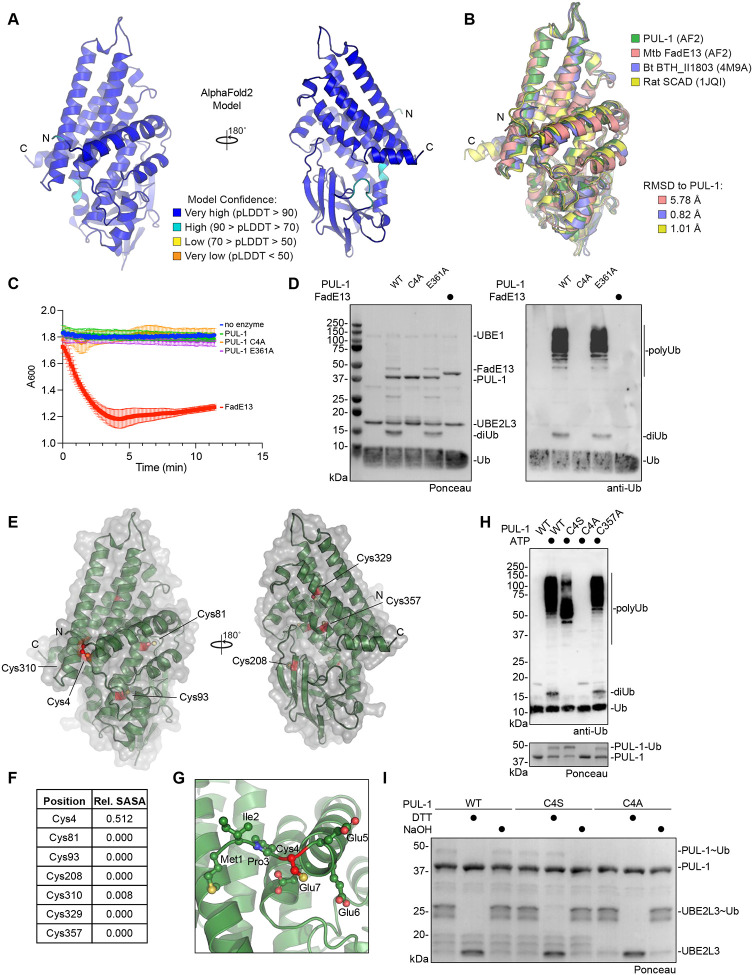
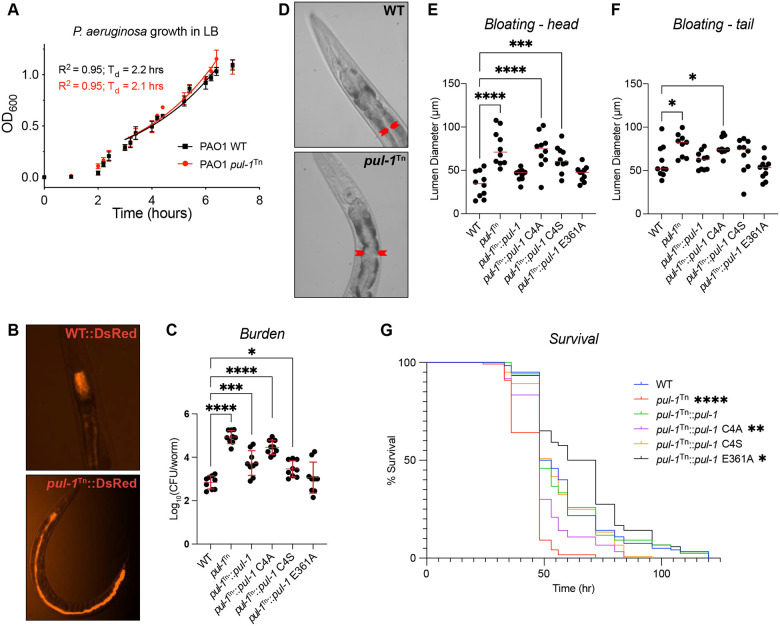
Ubiquitin signaling controls many aspects of eukaryotic biology, including targeted protein degradation and immune defense. Remarkably, invading bacterial pathogens have adapted secreted effector proteins that hijack host ubiquitination to gain control over host responses. These ubiquitin-targeted effectors can exhibit, for example, E3 ligase or deubiquitinase activities, often without any sequence or structural homology to eukaryotic ubiquitin regulators. Such convergence in function poses a challenge to the discovery of additional bacterial virulence factors that target ubiquitin. To overcome this, we have developed a workflow to harvest natively secreted bacterial effectors and functionally screen them for ubiquitin regulatory activities. After benchmarking this approach on diverse ligase and deubiquitinase activities from Salmonella Typhimurium, Enteropathogenic Escherichia coli, and Shigella flexneri, we applied it to the identification of a cryptic E3 ligase activity secreted by Pseudomonas aeruginosa. We identified an unreported P. aeruginosa E3 ligase, which we have termed Pseudomonas Ub ligase 1 (PUL-1), that resembles none of the other E3 ligases previously established in or outside of the eukaryotic system. Importantly, in an animal model of P. aeruginosa infection, PUL-1 ligase activity plays an important role in regulating virulence. Thus, our workflow for the functional identification of ubiquitin-targeted effector proteins carries promise for expanding our appreciation of how host ubiquitin regulation contributes to bacterial pathogenesis.
Keywords: Pseudomonas aeruginosa; Ubiquitin; bacterial effector; deubiquitinase; ubiquitin ligase.
Conflict of interest statement
COMPETING INTEREST STATEMENT The authors declare no competing interests.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources