

HuR (ELAVL1) regulates the CCHFV minigenome and HAZV replication by associating with viral genomic RNA
- PMID: 39348382
- PMCID: PMC11466401
- DOI: 10.1371/journal.pntd.0012553
HuR (ELAVL1) regulates the CCHFV minigenome and HAZV replication by associating with viral genomic RNA
Abstract
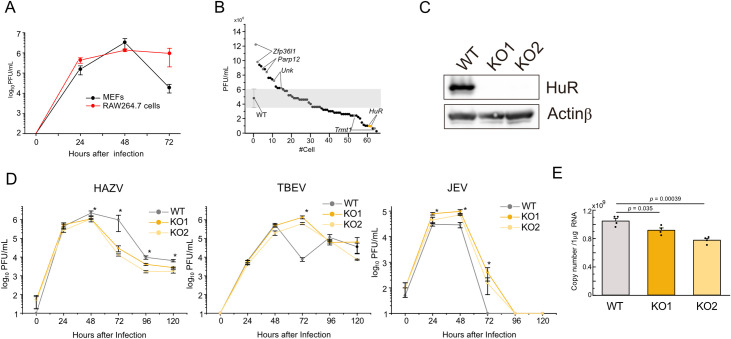
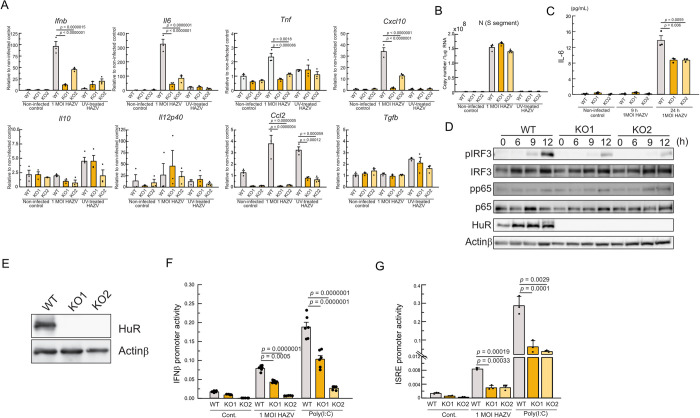
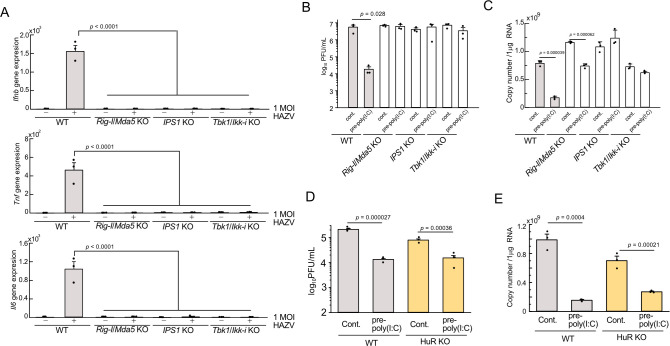
Crimean-Congo Hemorrhagic Fever virus (CCHFV) is a tick-borne pathogen that causes severe acute fever disease in humans and requires a biosafety level 4 laboratory for handling. Hazara virus (HAZV), belonging to the same virus genus as CCHFV, does not exhibit pathogenesis in humans. To investigate host RNA-binding proteins (RBPs) that regulate CCHFV replication, we generated a series of mutant RAW264.7 cells by CRISPR/Cas9 system and these cells were infected with HAZV. The viral titers in the supernatant of these cells was investigated, and HuR (ELAVL1) was identified. HuR KO RAW264.7 cells reduced HAZV replication. HuR is an RBP that enhances mRNA stability by binding to adenyl-uridine (AU)-rich regions in their 3' non-coding region (NCR). HuR regulates innate immune response by binding to host mRNAs of signaling molecules. The expression of cytokine genes such as Ifnb, Il6, and Tnf was reduced in HuR KO cells after HAZV infection. Although HuR supports the innate immune response during HAZV infection, we found that innate immune activation by HAZV infection did not affect its replication. We then investigated whether HuR regulates HAZV genome RNA stability. HAZV RNA genome was precipitated with an anti-HuR antibody, and HAZV genome RNA stability was lowered in HuR KO cells. We found that HuR associated with HAZV RNA and stabilized it to enhance HAZV replication. Furthermore, HuR-deficiency reduced CCHFV minigenome replication. CCHFV is a negative-strand RNA virus and positive-strand RNA is produced during replication. HuR was associated with positive-strand RNA rather than negative-strand RNA, and AU-rich region in 3'-NCR of S segment was responsible for immunoprecipitation with anti-HuR antibody and minigenome replication. Additionally, HuR inhibitor treatment reduced CCHFV minigenome replication. Our results indicate that HuR aids replication of the CCHFV minigenome by associating with the AU-rich region in the 3'-NCR.
Copyright: © 2024 Ikegawa et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
