

ATM1, an essential conserved transporter in Apicomplexa, bridges mitochondrial and cytosolic [Fe-S] biogenesis
- PMID: 39348385
- PMCID: PMC11476691
- DOI: 10.1371/journal.ppat.1012593
ATM1, an essential conserved transporter in Apicomplexa, bridges mitochondrial and cytosolic [Fe-S] biogenesis
Abstract
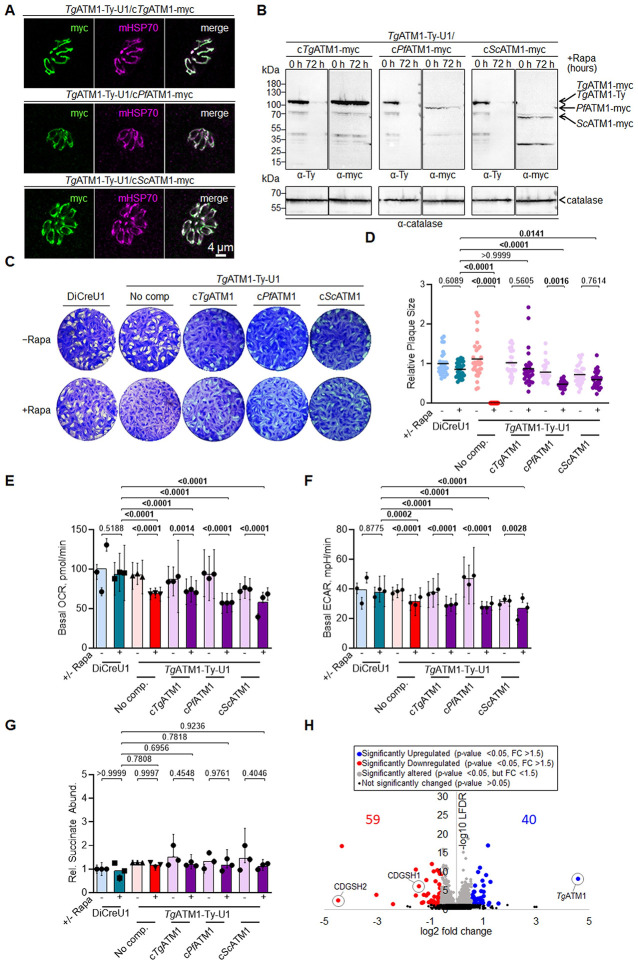
The Apicomplexa phylum encompasses numerous obligate intracellular parasites, some associated with severe implications for human health, including Plasmodium, Cryptosporidium, and Toxoplasma gondii. The iron-sulfur cluster [Fe-S] biogenesis ISC pathway, localized within the mitochondrion or mitosome of these parasites, is vital for parasite survival and development. Previous work on T. gondii and Plasmodium falciparum provided insights into the mechanisms of [Fe-S] biogenesis within this phylum, while the transporter linking mitochondria-generated [Fe-S] with the cytosolic [Fe-S] assembly (CIA) pathway remained elusive. This critical step is catalyzed by a well-conserved ABC transporter, termed ATM1 in yeast, ATM3 in plants and ABCB7 in mammals. Here, we identify and characterize this transporter in two clinically relevant Apicomplexa. We demonstrate that depletion of TgATM1 does not specifically impair mitochondrial metabolism. Instead, proteomic analyses reveal that TgATM1 expression levels inversely correlate with the abundance of proteins that participate in the transfer of [Fe-S] to cytosolic proteins at the outer mitochondrial membrane. Further insights into the role of TgATM1 are gained through functional complementation with the well-characterized yeast homolog. Biochemical characterization of PfATM1 confirms its role as a functional ABC transporter, modulated by oxidized glutathione (GSSG) and [4Fe-4S].
Copyright: © 2024 Shrivastava et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
