

Mycobacteria that cause tuberculosis have retained ancestrally acquired genes for the biosynthesis of chemically diverse terpene nucleosides
- PMID: 39348416
- PMCID: PMC11476799
- DOI: 10.1371/journal.pbio.3002813
Mycobacteria that cause tuberculosis have retained ancestrally acquired genes for the biosynthesis of chemically diverse terpene nucleosides
Abstract
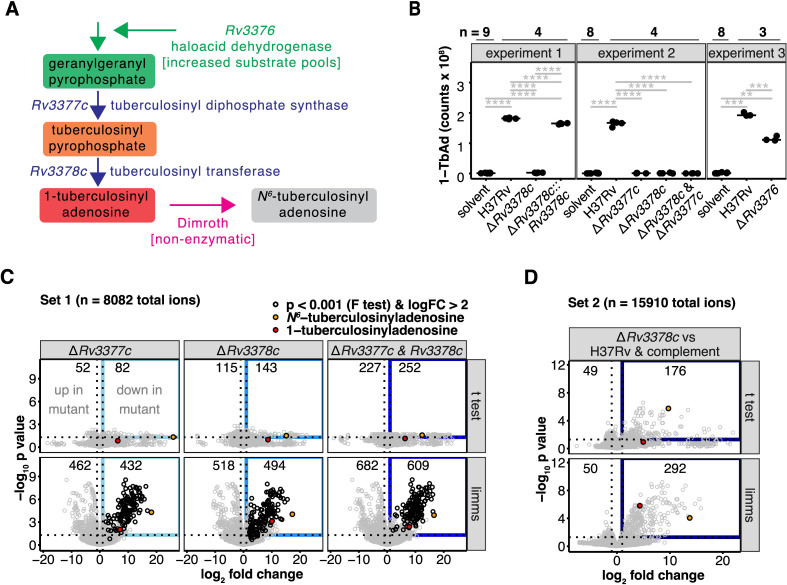
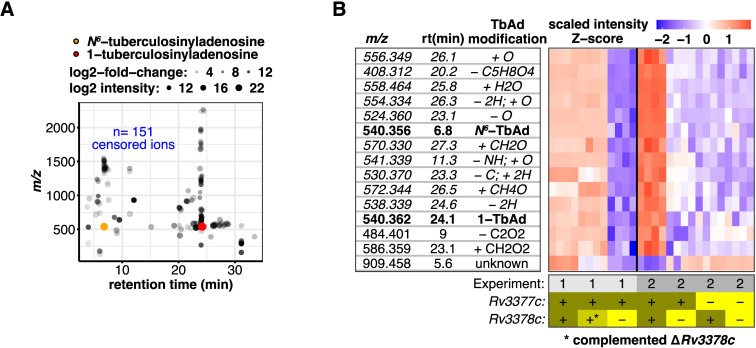
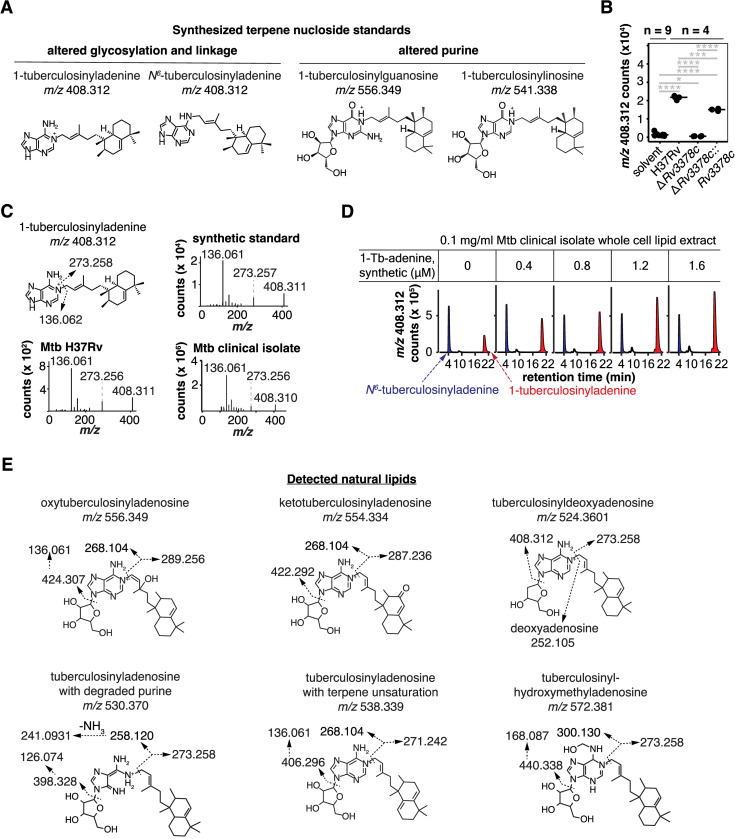
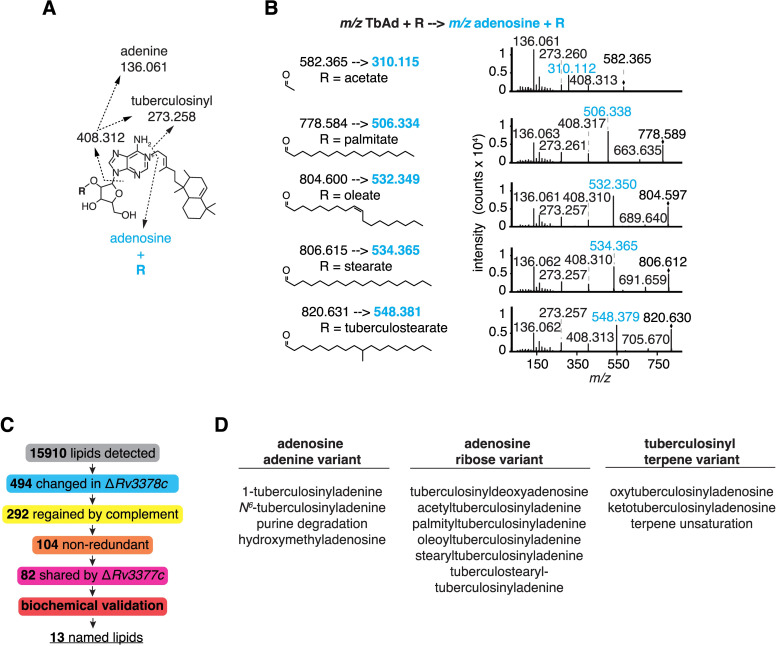
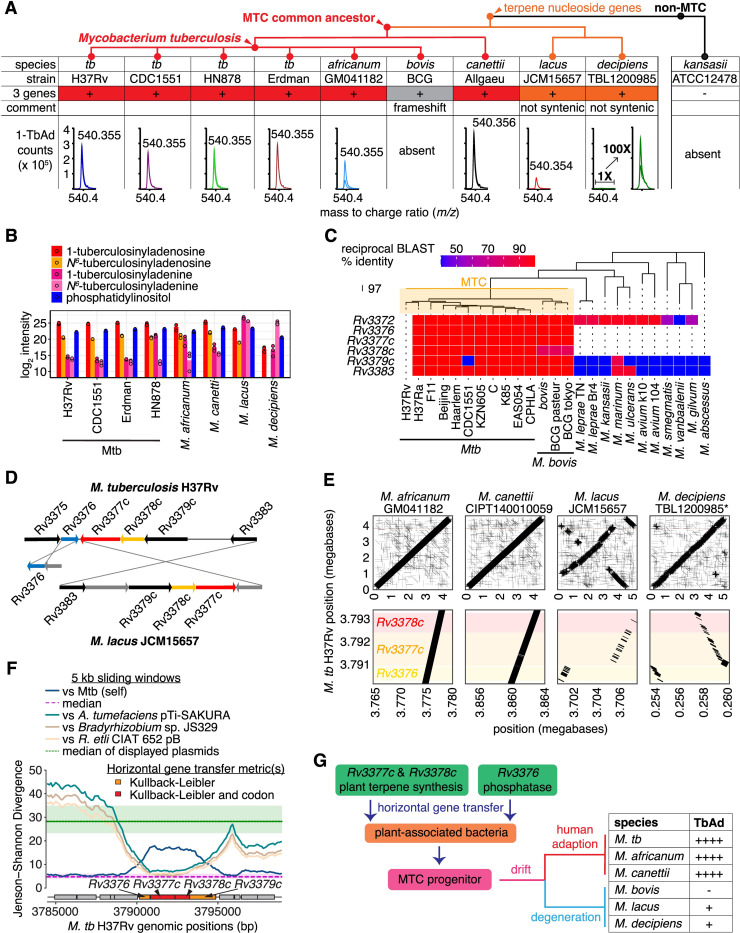
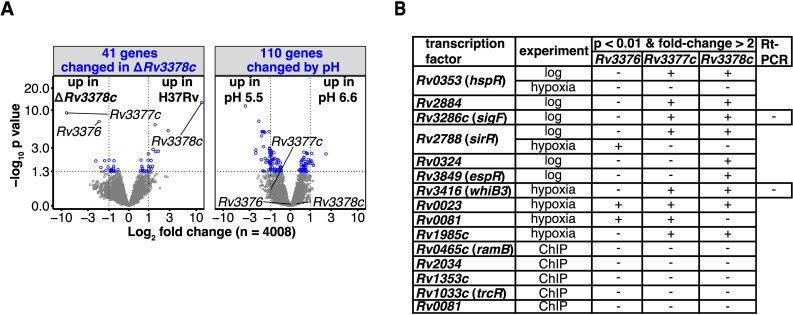
Mycobacterium tuberculosis (Mtb) releases the unusual terpene nucleoside 1-tuberculosinyladenosine (1-TbAd) to block lysosomal function and promote survival in human macrophages. Using conventional approaches, we found that genes Rv3377c and Rv3378c, but not Rv3376, were necessary for 1-TbAd biosynthesis. Here, we introduce linear models for mass spectrometry (limms) software as a next-generation lipidomics tool to study the essential functions of lipid biosynthetic enzymes on a whole-cell basis. Using limms, whole-cell lipid profiles deepened the phenotypic landscape of comparative mass spectrometry experiments and identified a large family of approximately 100 terpene nucleoside metabolites downstream of Rv3378c. We validated the identity of previously unknown adenine-, adenosine-, and lipid-modified tuberculosinol-containing molecules using synthetic chemistry and collisional mass spectrometry, including comprehensive profiling of bacterial lipids that fragment to adenine. We tracked terpene nucleoside genotypes and lipid phenotypes among Mycobacterium tuberculosis complex (MTC) species that did or did not evolve to productively infect either human or nonhuman mammals. Although 1-TbAd biosynthesis genes were thought to be restricted to the MTC, we identified the locus in unexpected species outside the MTC. Sequence analysis of the locus showed nucleotide usage characteristic of plasmids from plant-associated bacteria, clarifying the origin and timing of horizontal gene transfer to a pre-MTC progenitor. The data demonstrated correlation between high level terpene nucleoside biosynthesis and mycobacterial competence for human infection, and 2 mechanisms of 1-TbAd biosynthesis loss. Overall, the selective gain and evolutionary retention of tuberculosinyl metabolites in modern species that cause human TB suggest a role in human TB disease, and the newly discovered molecules represent candidate disease-specific biomarkers.
Copyright: © 2024 Mayfield et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Layre E, Lee HJ, Young DC, Martinot AJ, Buter J, Minnaard AJ, et al. Molecular profiling of Mycobacterium tuberculosis identifies tuberculosinyl nucleoside products of the virulence-associated enzyme Rv3378c. Proc Natl Acad Sci U S A. 2014;111(8):2978–83. doi: 10.1073/pnas.1315883111 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
