

Spatial interactions modulate tumor growth and immune infiltration
- PMID: 39349537
- PMCID: PMC11442770
- DOI: 10.1038/s41540-024-00438-1
Spatial interactions modulate tumor growth and immune infiltration
Abstract
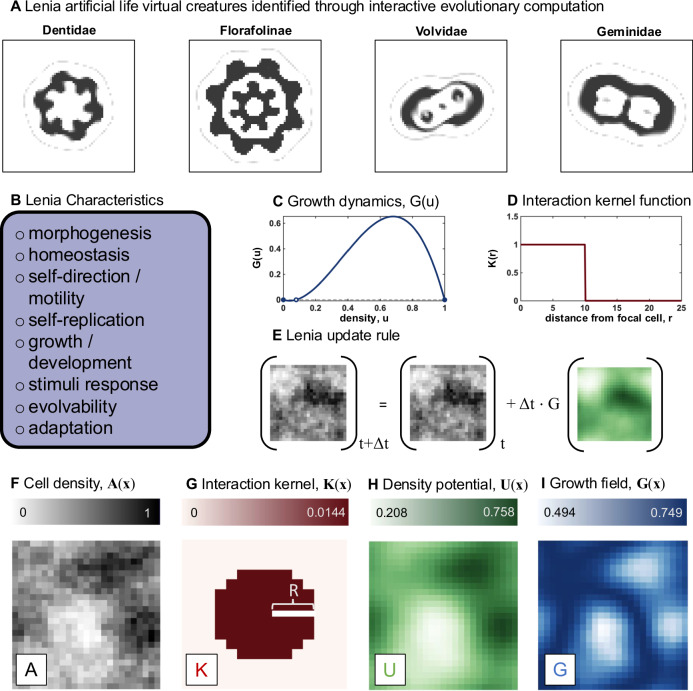
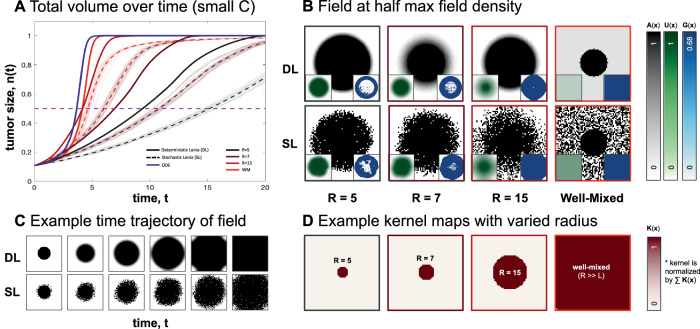
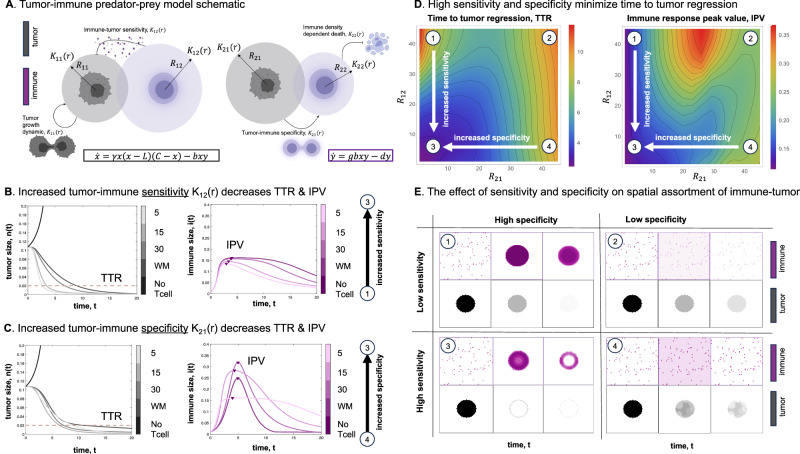
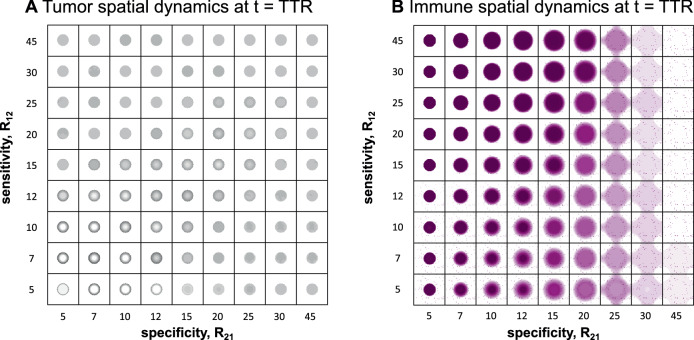
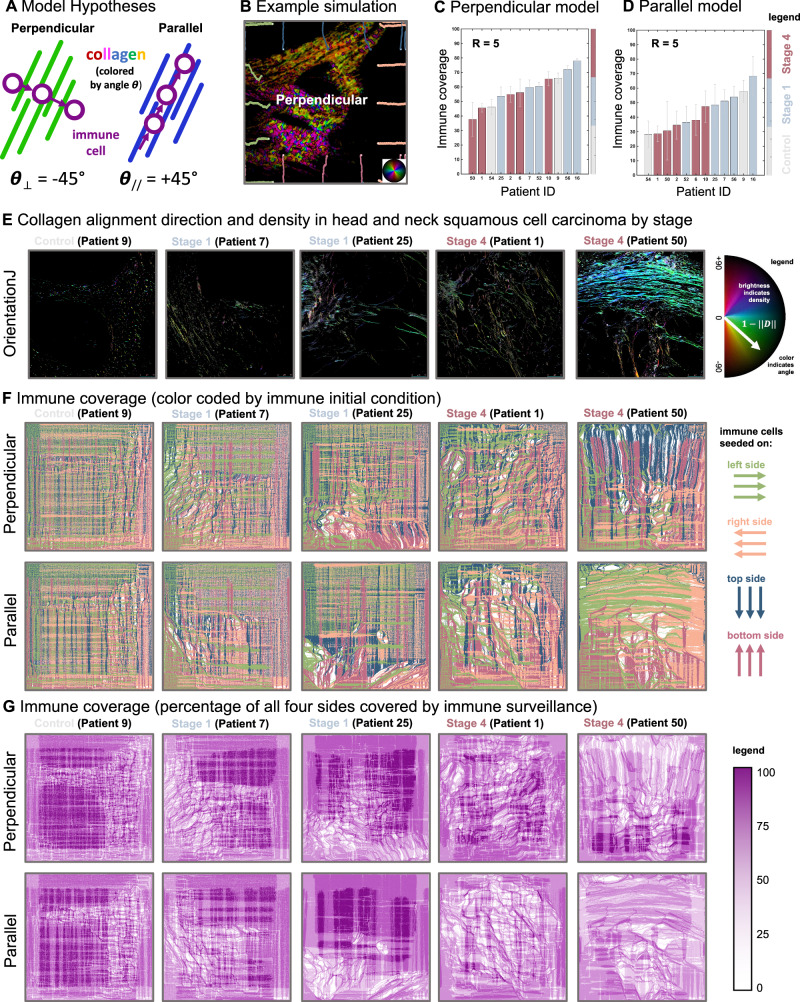
Direct observation of tumor-immune interactions is unlikely in tumors with currently available technology, but computational simulations based on clinical data can provide insight to test hypotheses. It is hypothesized that patterns of collagen evolve as a mechanism of immune escape, but the exact nature of immune-collagen interactions is poorly understood. Spatial data quantifying collagen fiber alignment in squamous cell carcinomas indicates that late-stage disease is associated with highly aligned fibers. Our computational modeling framework discriminates between two hypotheses: immune cell migration that moves (1) parallel or (2) perpendicular to collagen fiber orientation. The modeling recapitulates immune-extracellular matrix interactions where collagen patterns provide immune protection, leading to an emergent inverse relationship between disease stage and immune coverage. Here, computational modeling provides important mechanistic insights by defining a kernel cell-cell interaction function that considers a spectrum of local (cell-scale) to global (tumor-scale) spatial interactions. Short-range interaction kernels provide a mechanism for tumor cell survival under conditions with strong Allee effects, while asymmetric tumor-immune interaction kernels lead to poor immune response. Thus, the length scale of tumor-immune interaction kernels drives tumor growth and infiltration.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Update of
-
Spatial interactions modulate tumor growth and immune infiltration.bioRxiv [Preprint]. 2024 Mar 13:2024.01.10.575036. doi: 10.1101/2024.01.10.575036. bioRxiv. 2024. Update in: NPJ Syst Biol Appl. 2024 Sep 30;10(1):106. doi: 10.1038/s41540-024-00438-1. PMID: 38370722 Free PMC article. Updated. Preprint.
-
Spatial interactions modulate tumor growth and immune infiltration.Res Sq [Preprint]. 2024 May 22:rs.3.rs-3962451. doi: 10.21203/rs.3.rs-3962451/v1. Res Sq. 2024. Update in: NPJ Syst Biol Appl. 2024 Sep 30;10(1):106. doi: 10.1038/s41540-024-00438-1. PMID: 38826398 Free PMC article. Updated. Preprint.
References
-
- Chan, B. W.-C. Lenia-biology of artificial life. Complex Syst. 28, 251–286 (2019).
-
- Chan, B. W.-C. Lenia and expanded universe. In: Artificial Life Conference Proceedings 32, 221–229 (2020).
-
- Jain, S. & Shrestha, A. Capturing emerging complexity in Lenia. arXivhttps://arxiv.org/abs/2305.09378 (2023).
-
- Gardner, M. Mathematical games-the fantastic combinations of john conway’s new solitaire game, life, 1970. Sci. Am.223, 120–123 (1970).
-
- Davis, Q. T. & Bongard, J. Glaberish: generalizing the continuously-valued Lenia framework to arbitrary life-like cellular automata. In: Artificial Life Conference Proceedings 34, 2022, 47 (2022).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
