

The IgLON family of cell adhesion molecules expressed in developing neural circuits ensure the proper functioning of the sensory system in mice
- PMID: 39349721
- PMCID: PMC11442611
- DOI: 10.1038/s41598-024-73358-z
The IgLON family of cell adhesion molecules expressed in developing neural circuits ensure the proper functioning of the sensory system in mice
Abstract
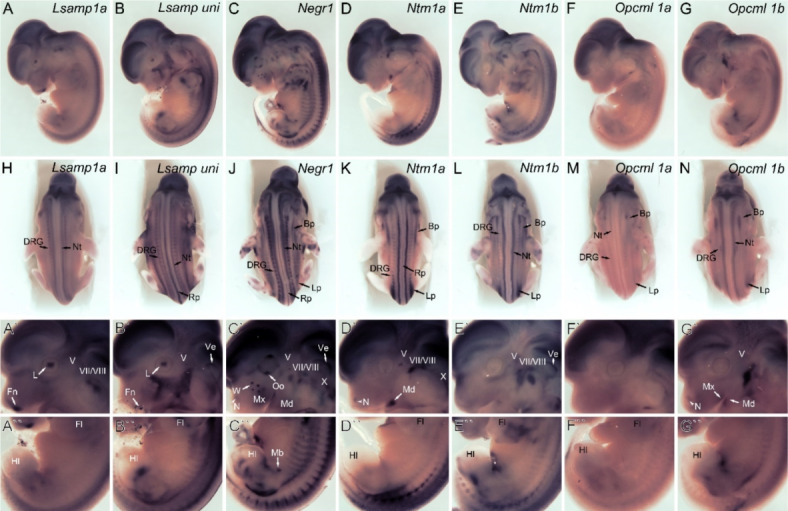
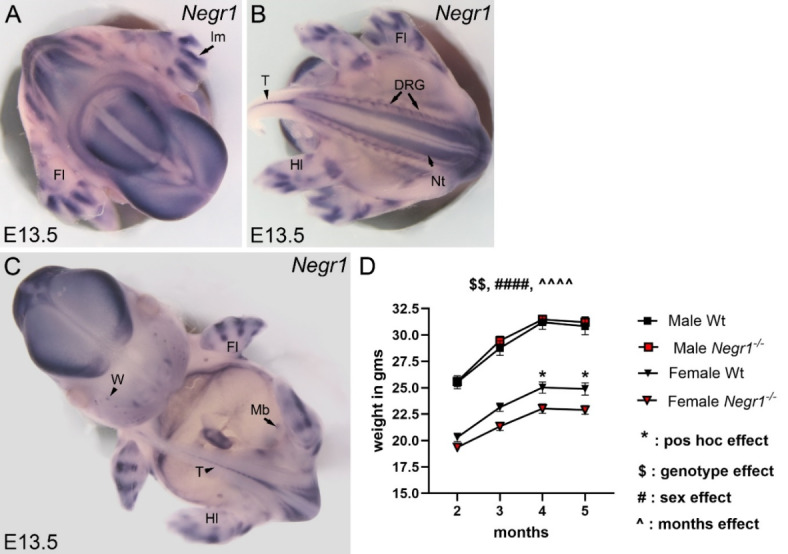
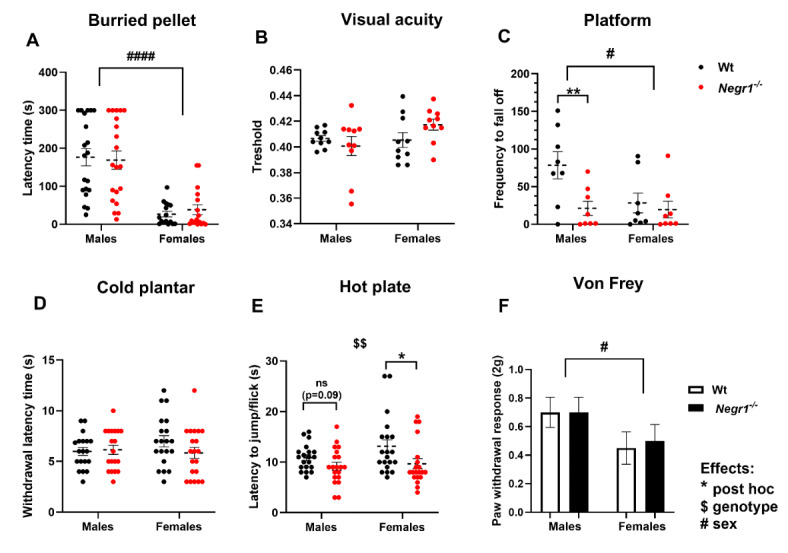
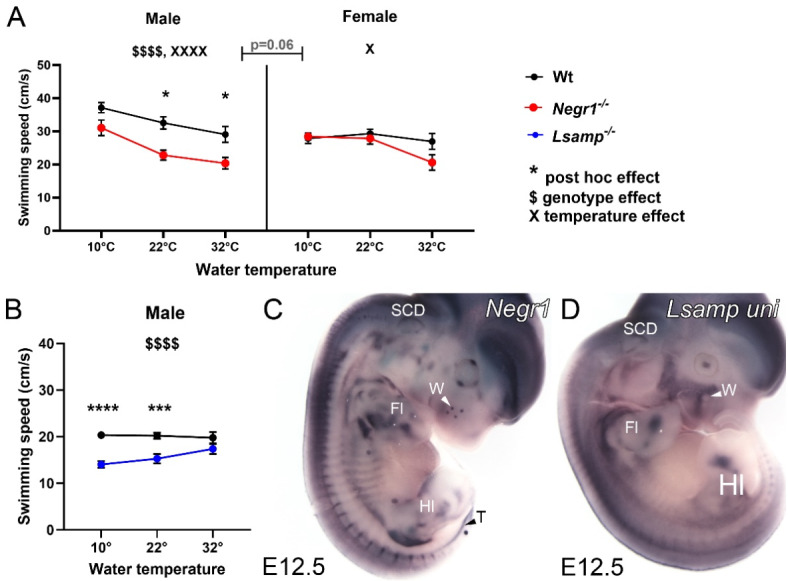
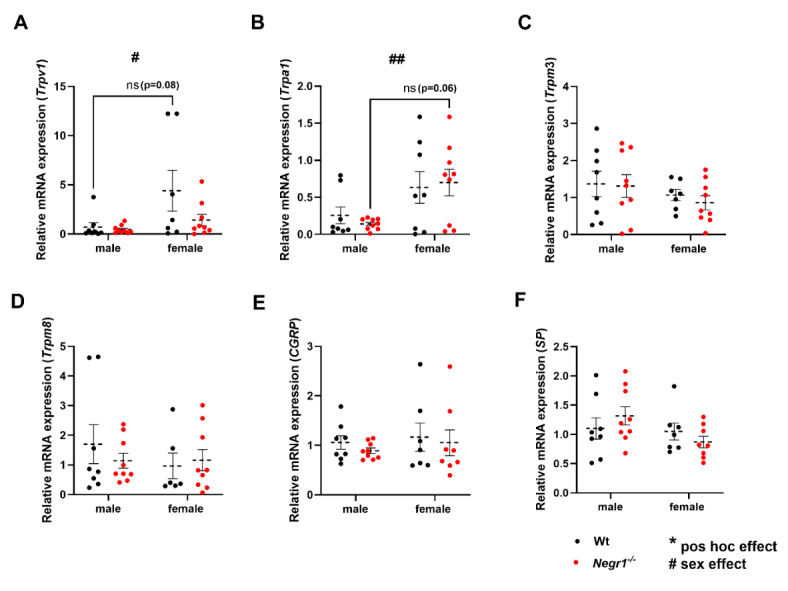
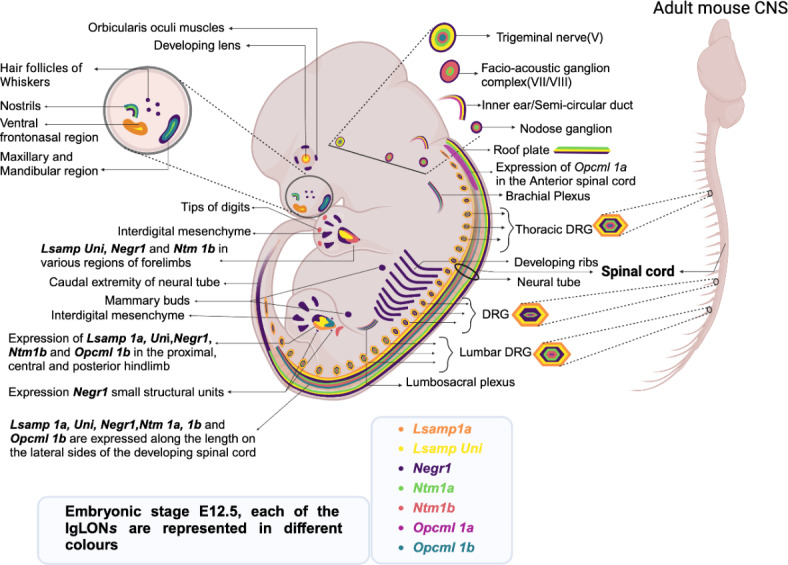
Deletions and malfunctions of the IgLON family of cell adhesion molecules are associated with anatomical, behavioral, and metabolic manifestations of neuropsychiatric disorders. We have previously shown that IgLON genes are expressed in sensory nuclei/pathways and that IgLON proteins modulate sensory processing. Here, we examined the expression of IgLON alternative promoter-specific isoforms during embryonic development and studied the sensory consequences of the anatomical changes when one of the IgLON genes, Negr1, is knocked out. At the embryonal age of E12.5 and E13.5, various IgLONs were distributed differentially and dynamically in the developing sensory areas within the central and peripheral nervous system, as well as in limbs and mammary glands. Sensory tests showed that Negr1 deficiency causes differences in vestibular function and temperature sensitivity in the knockout mice. Sex-specific differences were noted across olfaction, vestibular functioning, temperature regulation, and mechanical sensitivity. Our findings highlight the involvement of IgLON molecules during sensory circuit formation and suggest Negr1's critical role in somatosensory processing.
Keywords: DRG; IgLON; LSAMP; NEGR1; PNS; Sensory processing.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Catala, M. & Kubis, N. Gross anatomy and development of the peripheral nervous system. Handb. Clin. Neurol.115, 29–41. 10.1016/B978-0-444-52902-2.00003-5 (2013). - PubMed
-
- Baum, M. J. & Cherry, J. A. Processing by the main olfactory system of chemosignals that facilitate mammalian reproduction. Horm. Behav.68, 53–64. 10.1016/j.yhbeh.2014.06.003 (2015). - PubMed
-
- Burke, K. & Kobrina, A. Rodentia sensory systems. In Encyclopedia of Animal Cognition and Behavior (eds Vonk, J. & Shackelford, T. K.) (Springer, 2022). 10.1007/978-3-319-55065-7_765.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
