

Rapid and local neuroestrogen synthesis supports long-term potentiation of hippocampal Schaffer collaterals-cornu ammonis 1 synapse in ovariectomized mice
- PMID: 39351868
- PMCID: PMC11646664
- DOI: 10.1111/jne.13450
Rapid and local neuroestrogen synthesis supports long-term potentiation of hippocampal Schaffer collaterals-cornu ammonis 1 synapse in ovariectomized mice
Abstract
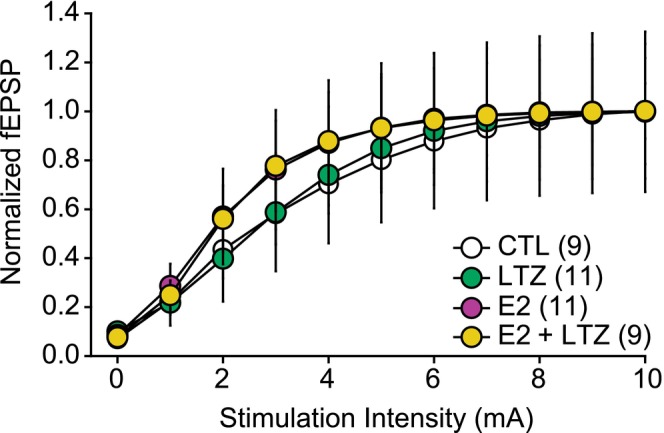
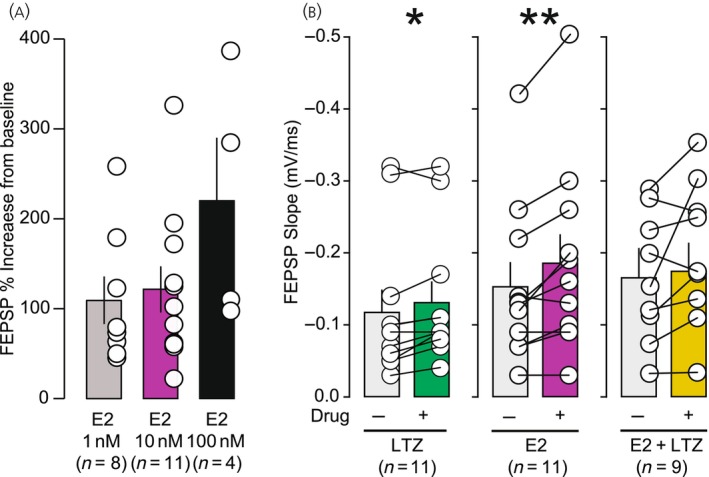
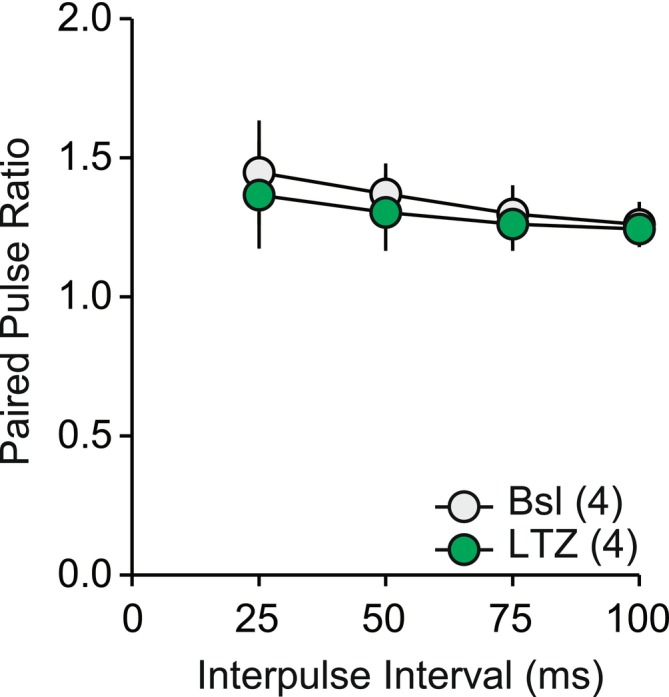
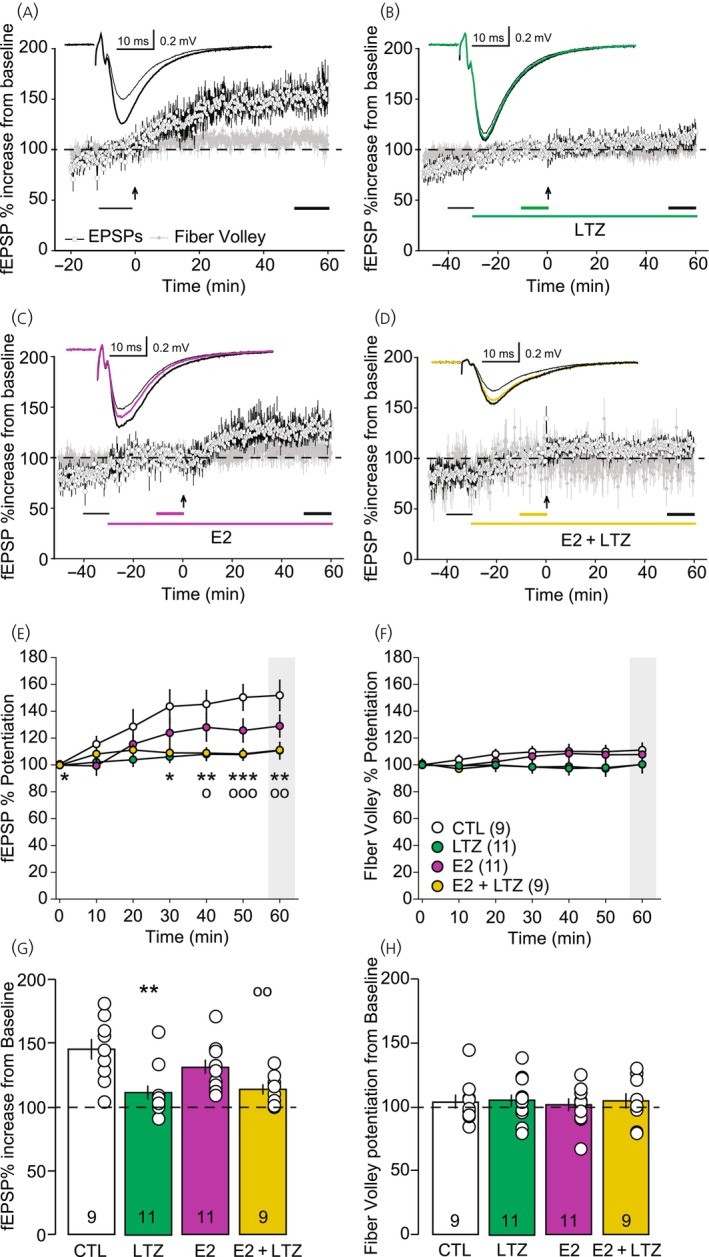
In aging women, cognitive decline and increased risk of dementia have been associated with the cessation of ovarian hormones production at menopause. In the brain, presence of the key enzyme aromatase required for the synthesis of 17-β-estradiol (E2) allows for local production of E2 in absence of functional ovaries. Understanding how aromatase activity is regulated could help alleviate the cognitive symptoms. In female rodents, genetic or pharmacological reduction of aromatase activity over extended periods of time impair memory formation, decreases spine density, and hinders long-term potentiation (LTP) in the hippocampus. Conversely, increased excitatory neurotransmission resulting in rapid N-methyl-d-aspartic acid (NMDA) receptor activation rapidly promotes neuroestrogen synthesis. This rapid modulation of aromatase activity led us to address the hypothesis that acute neuroestrogens synthesis is necessary for LTP at the Schaffer collateral-cornu ammonis 1 (CA1) synapse in absence of circulating ovarian estrogens. To test this hypothesis, we did electrophysiological recordings of field excitatory postsynaptic potential (fEPSPs) in hippocampal slices obtained from ovariectomized mice. To assess the impact of neuroestrogens synthesis on LTP, we applied the specific aromatase inhibitor, letrozole, before the induction of LTP with a theta burst stimulation protocol. We found that blocking aromatase activity prevented LTP. Interestingly, exogenous E2 application, while blocking aromatase activity, was not sufficient to recover LTP in our model. Our results indicate the critical importance of rapid, activity-dependent local neuroestrogens synthesis, independent of circulating hormones for hippocampal synaptic plasticity in female rodents.
Keywords: aromatase; electrophysiology; estrogen; hippocampus; synaptic plasticity.
© 2024 The Author(s). Journal of Neuroendocrinology published by John Wiley & Sons Ltd on behalf of British Society for Neuroendocrinology.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Pines A. Alzheimer's disease, menopause and the impact of the estrogenic environment. Climacteric. 2016;19:430‐432. - PubMed
-
- Daniel JM, Hulst JL, Berbling JL. Estradiol replacement enhances working memory in middle‐aged rats when initiated immediately after Ovariectomy but not after a long‐term period of ovarian hormone deprivation. Endocrinology. 2006;147:607‐614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
