

Chronically Low NMNAT2 Expression Causes Sub-lethal SARM1 Activation and Altered Response to Nicotinamide Riboside in Axons
- PMID: 39352636
- PMCID: PMC11790816
- DOI: 10.1007/s12035-024-04480-2
Chronically Low NMNAT2 Expression Causes Sub-lethal SARM1 Activation and Altered Response to Nicotinamide Riboside in Axons
Abstract
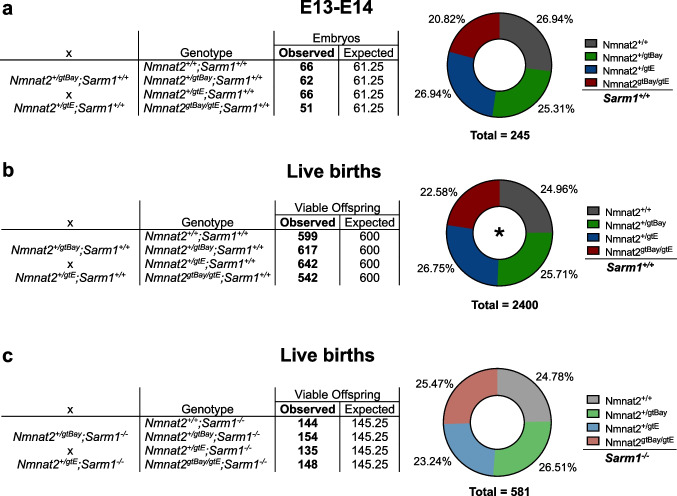
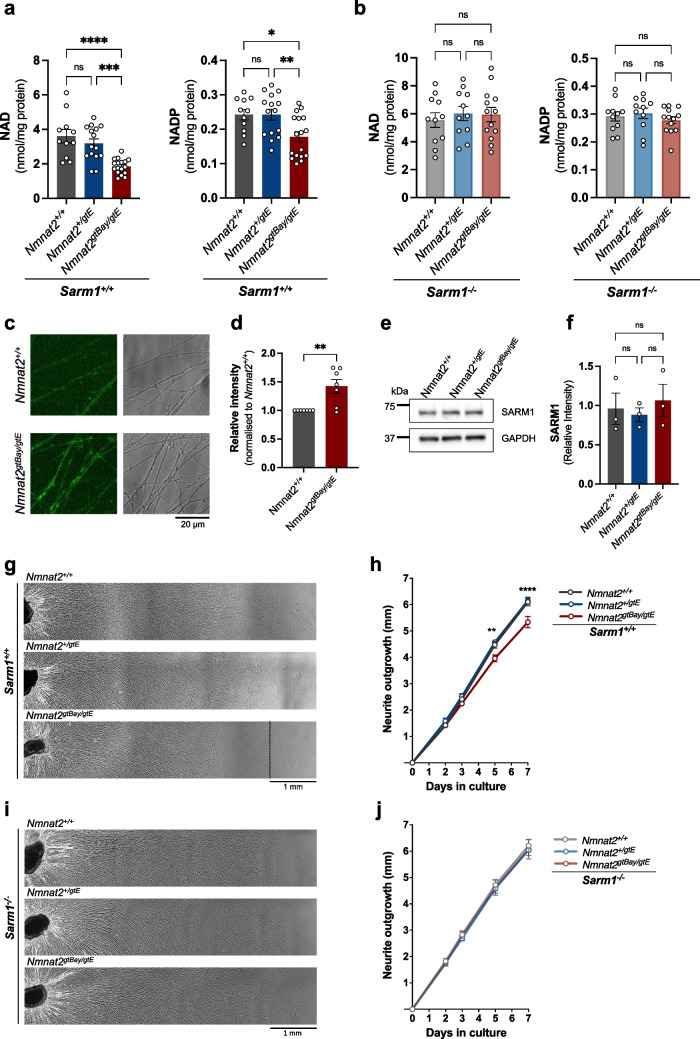
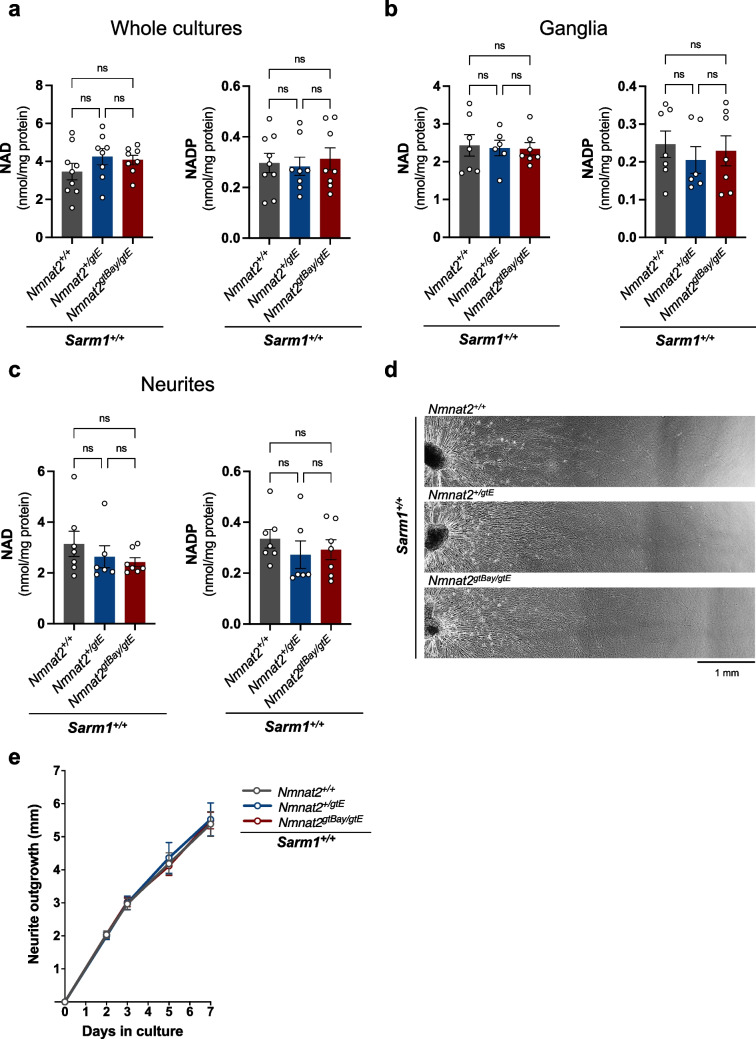
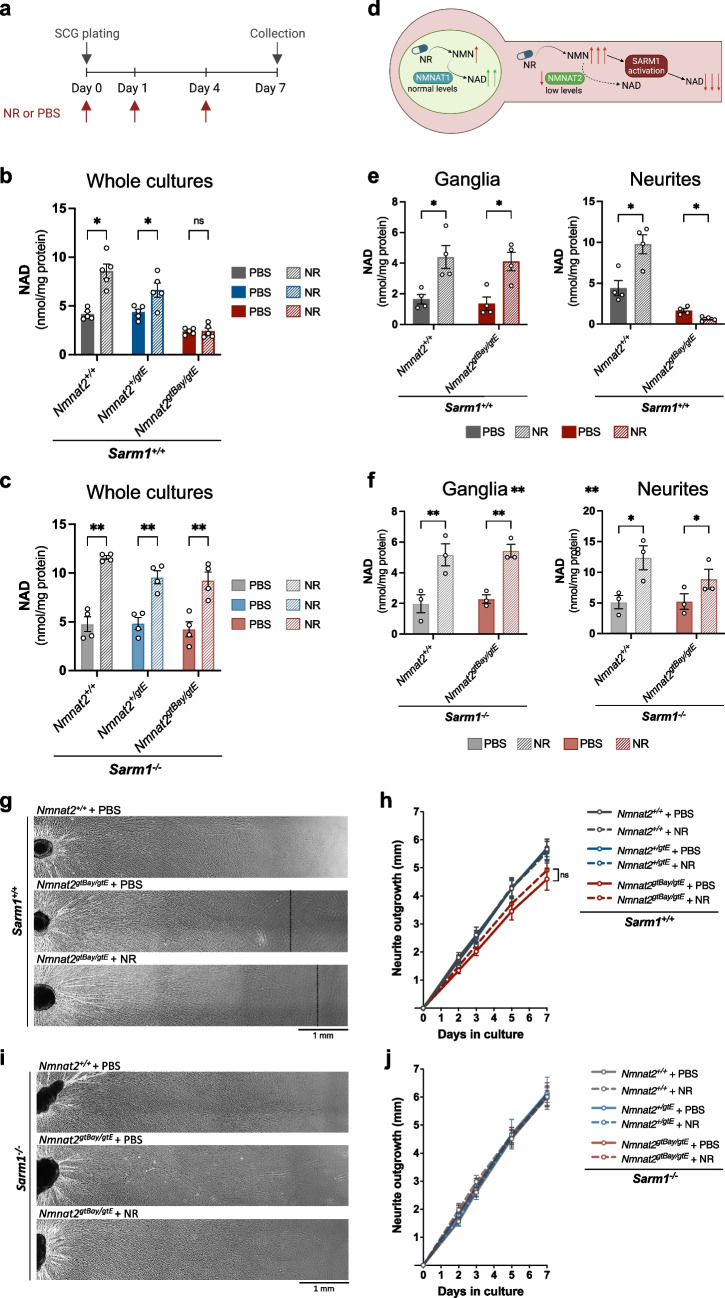
Nicotinamide mononucleotide adenylyltransferase 2 (NMNAT2) is an endogenous axon survival factor that maintains axon health by blocking activation of the downstream pro-degenerative protein SARM1 (sterile alpha and TIR motif containing protein 1). While complete absence of NMNAT2 in mice results in extensive axon truncation and perinatal lethality, the removal of SARM1 completely rescues these phenotypes. Reduced levels of NMNAT2 can be compatible with life; however, they compromise axon development and survival. Mice born expressing sub-heterozygous levels of NMNAT2 remain overtly normal into old age but develop axonal defects in vivo and in vitro as well as behavioural phenotypes. Therefore, it is important to examine the effects of constitutively low NMNAT2 expression on SARM1 activation and disease susceptibility. Here we demonstrate that chronically low NMNAT2 levels reduce prenatal viability in mice in a SARM1-dependent manner and lead to sub-lethal SARM1 activation in morphologically intact axons of superior cervical ganglion (SCG) primary cultures. This is characterised by a depletion in NAD(P) and compromised neurite outgrowth. We also show that chronically low NMNAT2 expression reverses the NAD-enhancing effect of nicotinamide riboside (NR) in axons in a SARM1-dependent manner. These data indicate that low NMNAT2 levels can trigger sub-lethal SARM1 activation which is detectable at the molecular level and could predispose to human axonal disorders.
Keywords: NAD; NMNAT2; Programmed axon death; SARM1.
© 2024. The Author(s).
Conflict of interest statement
Declarations. Ethics Approval: Animal work was approved by the University of Cambridge and performed in accordance with the Home Office Animal Scientific Procedures Act (ASPA), 1986 under project licence P98A03BF9. Consent to Participate: Not applicable. Consent for Publication: Not applicable. Competing Interests: MPC consults for Nura Bio and Drishti Discoveries and the Coleman group is part funded by AstraZeneca for academic research projects but none of these activities relate to the study reported here.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
