

Angiogenesis is limited by LIC1-mediated lysosomal trafficking
- PMID: 39356418
- PMCID: PMC11653708
- DOI: 10.1007/s10456-024-09951-7
Angiogenesis is limited by LIC1-mediated lysosomal trafficking
Abstract
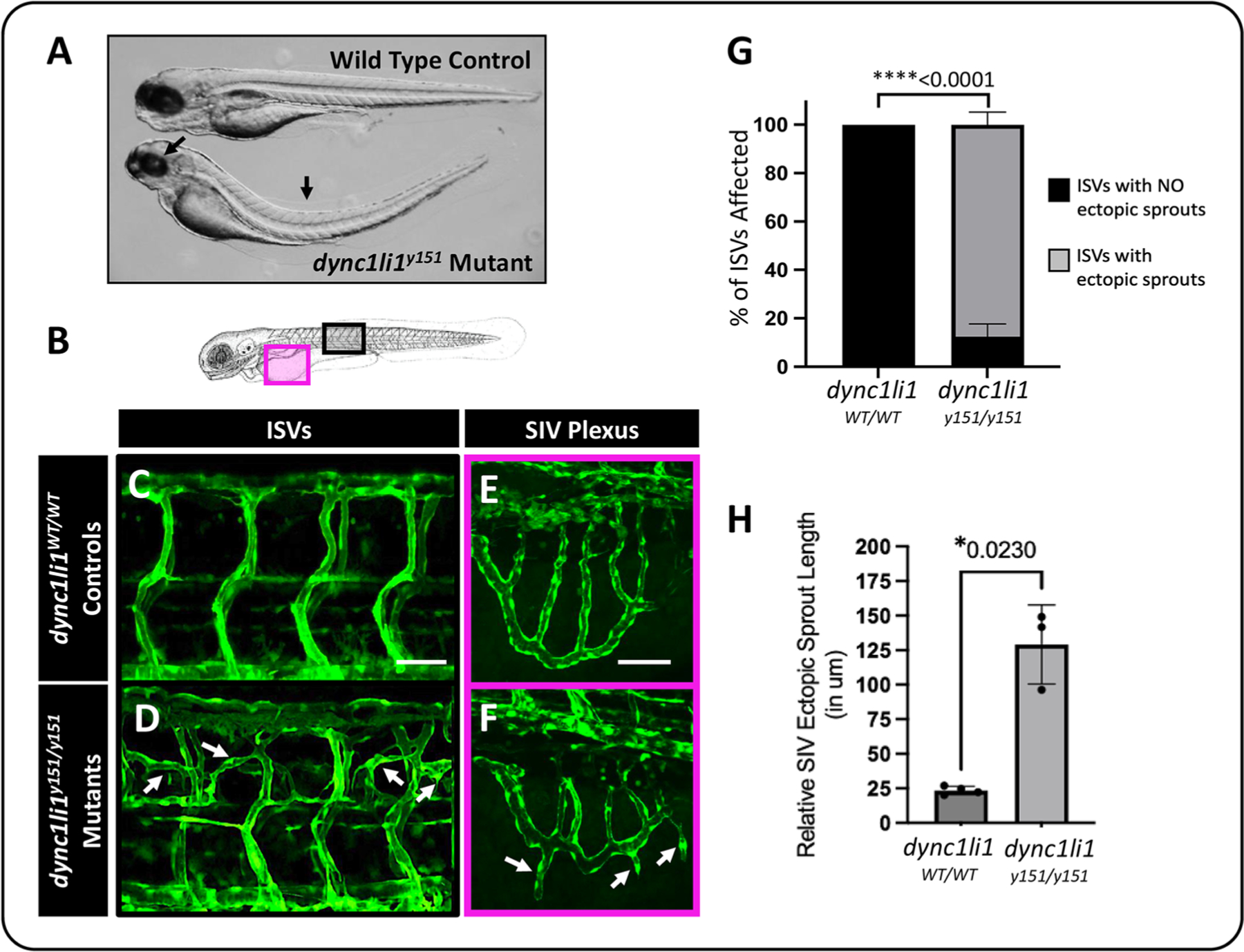
Dynein cytoplasmic 1 light intermediate chain 1 (LIC1, DYNC1LI1) is a core subunit of the dynein motor complex. The LIC1 subunit also interacts with various cargo adaptors to regulate Rab-mediated endosomal recycling and lysosomal degradation. Defects in this gene are predicted to alter dynein motor function, Rab binding capabilities, and cytoplasmic cargo trafficking. Here, we have identified a dync1li1 zebrafish mutant, harboring a premature stop codon at the exon 12/13 splice acceptor site, that displays increased angiogenesis. In vitro, LIC1-deficient human endothelial cells display increases in cell surface levels of the pro-angiogenic receptor VEGFR2, SRC phosphorylation, and Rab11-mediated endosomal recycling. In vivo, endothelial-specific expression of constitutively active Rab11a leads to excessive angiogenesis, similar to the dync1li1 mutants. Increased angiogenesis is also evident in zebrafish harboring mutations in rilpl1/2, the adaptor proteins that promote Rab docking to Lic1 to mediate lysosomal targeting. These findings suggest that LIC1 and the Rab-adaptor proteins RILPL1 and 2 restrict angiogenesis by promoting degradation of VEGFR2-containing recycling endosomes. Disruption of LIC1- and RILPL1/2-mediated lysosomal targeting increases Rab11-mediated recycling endosome activity, promoting excessive SRC signaling and angiogenesis.
Keywords: Angiogenesis; Dynein motor; Endosomes; Lic1; Lysosomes; Rilp1/2; Zebrafish.
© 2024. The Author(s), under exclusive licence to Springer Nature B.V.
Conflict of interest statement
Figures







Update of
-
Regulation of angiogenesis by endocytic trafficking mediated by cytoplasmic dynein 1 light intermediate chain 1.bioRxiv [Preprint]. 2024 Apr 1:2024.04.01.587559. doi: 10.1101/2024.04.01.587559. bioRxiv. 2024. Update in: Angiogenesis. 2024 Nov;27(4):943-962. doi: 10.1007/s10456-024-09951-7. PMID: 38903077 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- ZIA HD001607/ImNIH/Intramural NIH HHS/United States
- ZIA HD001011/ImNIH/Intramural NIH HHS/United States
- ZIA HD001011-27/Intramural Program of NICHD
- R35GM142779/NIH/NIGMS
- K99 HL171944/HL/NHLBI NIH HHS/United States
- R35GM137976/NIH/NIGMS
- PRE/American Heart Association Predoctoral Fellowship
- UL1 TR002345/TR/NCATS NIH HHS/United States
- K99HL171944/NIH/NHLBI
- R35 GM142779/GM/NIGMS NIH HHS/United States
- R35 GM137976/GM/NIGMS NIH HHS/United States
- UL1TR002345/Washington University Institute of Clinical and Translational Sciences Which Is, in Part, Supported by the NIH/National Center for Advancing Translational Sciences (NCATS)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
