

Pachytene piRNAs control discrete meiotic events during spermatogenesis and restrict gene expression in space and time
- PMID: 39356768
- PMCID: PMC11446278
- DOI: 10.1126/sciadv.adp0466
Pachytene piRNAs control discrete meiotic events during spermatogenesis and restrict gene expression in space and time
Abstract
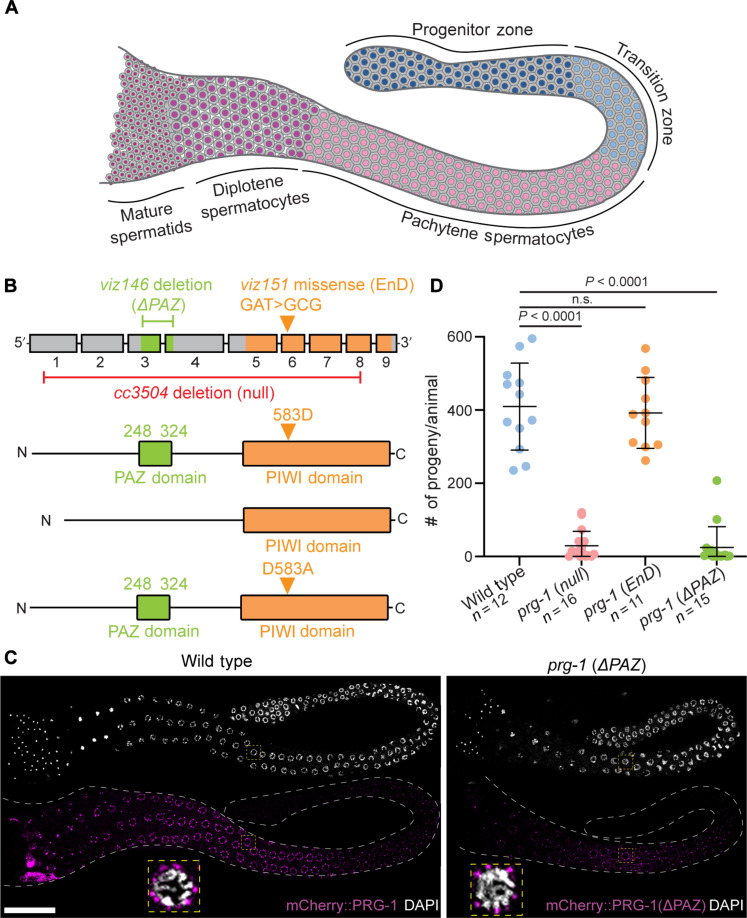
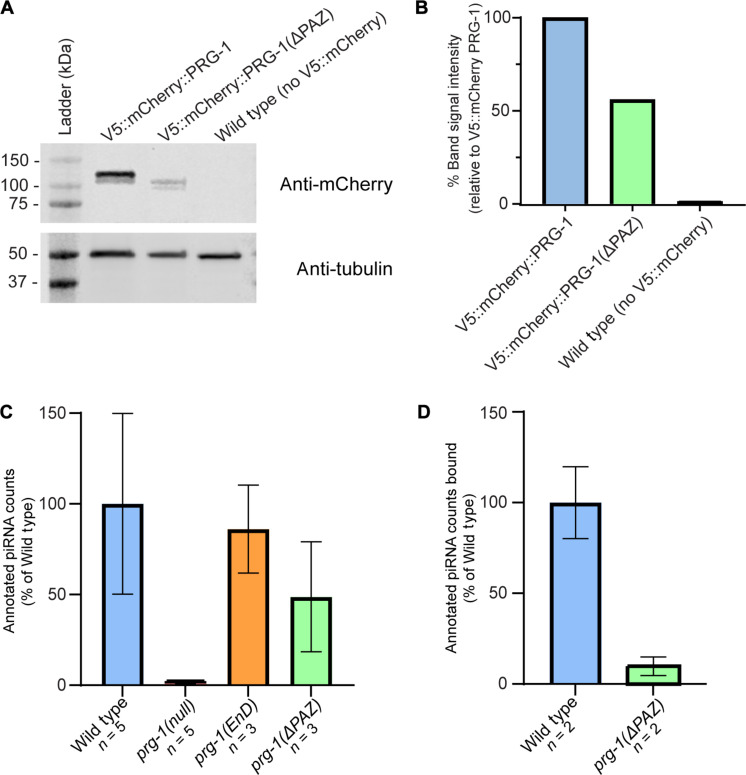
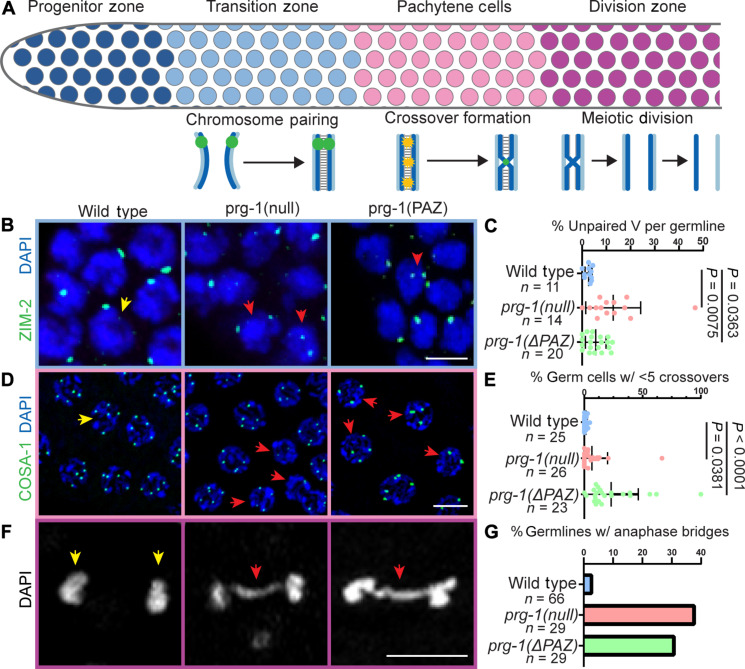
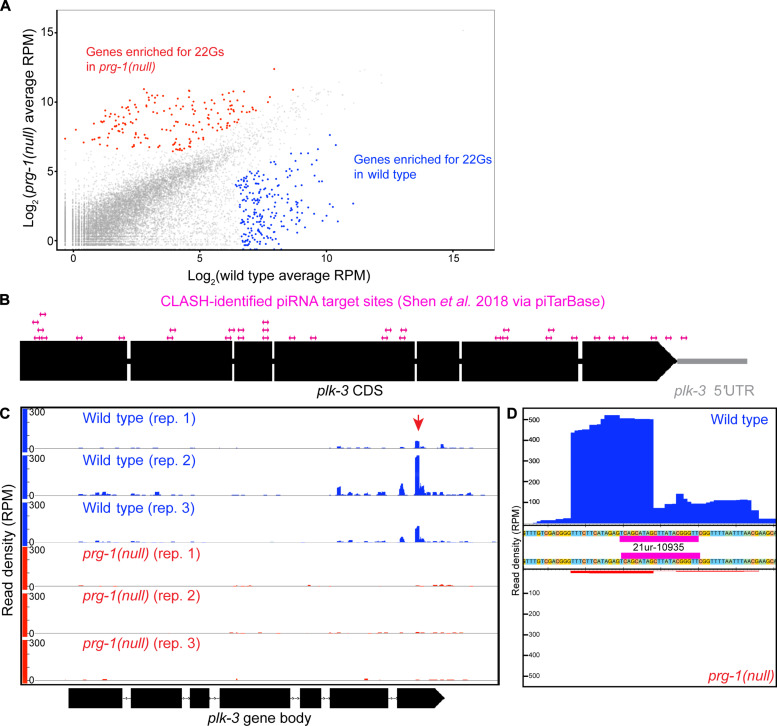
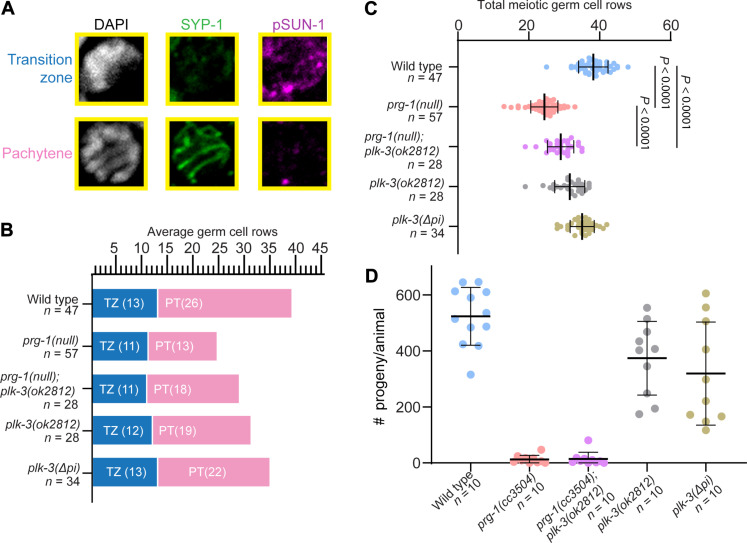
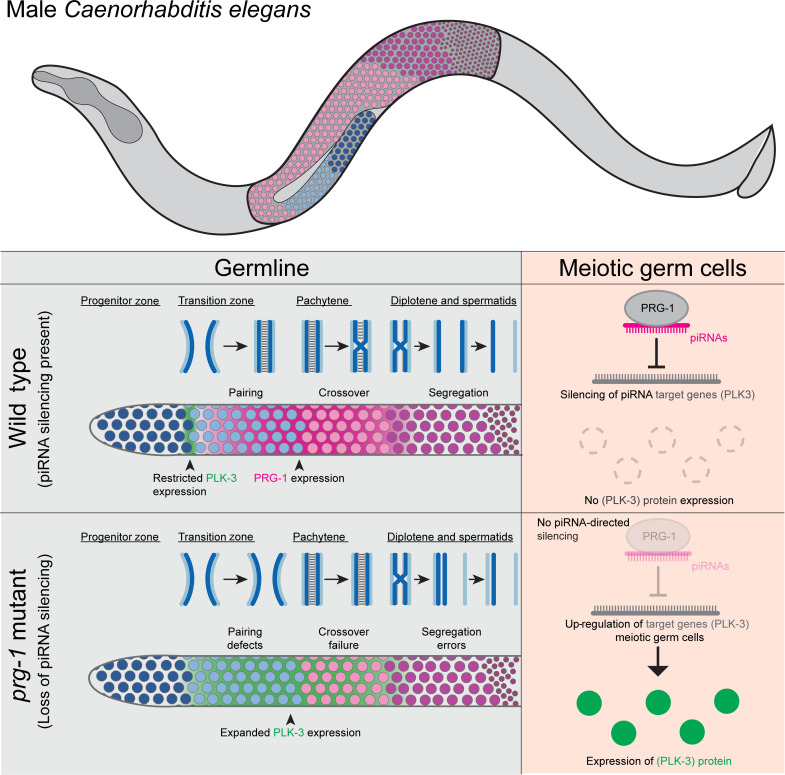
Pachytene piRNAs, a Piwi-interacting RNA subclass in mammals, are hypothesized to regulate non-transposon sequences during spermatogenesis. Caenorhabditis elegans piRNAs, the 21URNAs, are implicated in regulating coding sequences; the messenger RNA targets and biological processes they control during spermatogenesis are largely unknown. We demonstrate that loss of 21URNAs compromises homolog pairing and makes it permissive for nonhomologous synapsis resulting in defects in crossover formation and chromosome segregation during spermatogenesis. We identify Polo-like kinase 3 (PLK-3), among others, as a 21URNA target. 21URNA activity restricts PLK-3 protein to proliferative cells, and expansion of PLK-3 in pachytene overlaps with the meiotic defects. Removal of plk-3 results in quantitative genetic suppression of the meiotic defects. One discrete 21URNA inhibits PLK-3 expression in late pachytene cells. Together, these results suggest that the 21URNAs function as pachytene piRNAs during C. elegans spermatogenesis. We identify their targets and meiotic events and highlight the remarkable intricacy of this multi-effector mechanism during spermatogenesis.
Figures

References
-
- Aravin A., Gaidatzis D., Pfeffer S., Lagos-Quintana M., Landgraf P., Iovino N., Morris P., Brownstein M. J., Kuramochi-Miyagawa S., Nakano T., Chien M., Russo J. J., Ju J., Sheridan R., Sander C., Zavolan M., Tuschl T., A novel class of small RNAs bind to MILI protein in mouse testes. Nature 442, 203–207 (2006). - PubMed
-
- Vagin V. V., Sigova A., Li C., Seitz H., Gvozdev V., Zamore P. D., A distinct small RNA pathway silences selfish genetic elements in the germline. Science 313, 320–324 (2006). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
