

Inertial effect of cell state velocity on the quiescence-proliferation fate decision
- PMID: 39358384
- PMCID: PMC11447052
- DOI: 10.1038/s41540-024-00428-3
Inertial effect of cell state velocity on the quiescence-proliferation fate decision
Abstract
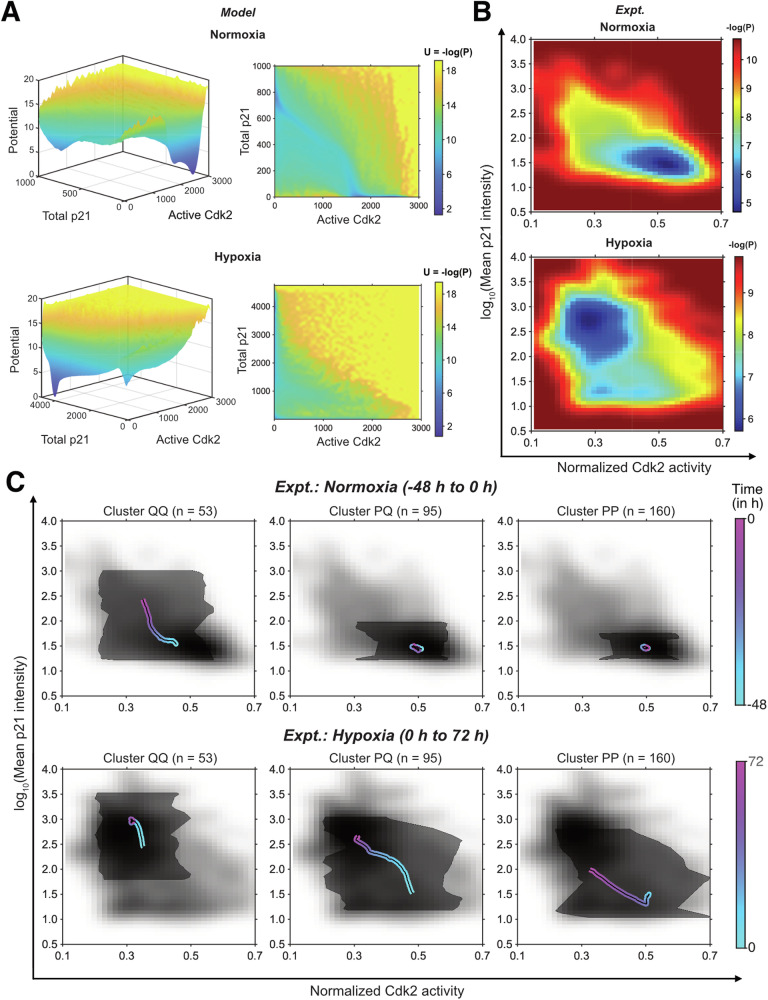
Energy landscapes can provide intuitive depictions of population heterogeneity and dynamics. However, it is unclear whether individual cell behavior, hypothesized to be determined by initial position and noise, is faithfully recapitulated. Using the p21-/Cdk2-dependent quiescence-proliferation decision in breast cancer dormancy as a testbed, we examined single-cell dynamics on the landscape when perturbed by hypoxia, a dormancy-inducing stress. Combining trajectory-based energy landscape generation with single-cell time-lapse microscopy, we found that a combination of initial position and velocity on a p21/Cdk2 landscape, but not position alone, was required to explain the observed cell fate heterogeneity under hypoxia. This is likely due to additional cell state information such as epigenetic features and/or other species encoded in velocity but missing in instantaneous position determined by p21 and Cdk2 levels alone. Here, velocity dependence manifested as inertia: cells with higher cell cycle velocities prior to hypoxia continued progressing along the cell cycle under hypoxia, resisting the change in landscape towards cell cycle exit. Such inertial effects may markedly influence cell fate trajectories in tumors and other dynamically changing microenvironments where cell state transitions are governed by coordination across several biochemical species.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Update of
-
Inertial effect of cell state velocity on the quiescence-proliferation fate decision in breast cancer.bioRxiv [Preprint]. 2023 May 24:2023.05.22.541793. doi: 10.1101/2023.05.22.541793. bioRxiv. 2023. Update in: NPJ Syst Biol Appl. 2024 Oct 2;10(1):111. doi: 10.1038/s41540-024-00428-3. PMID: 37292599 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
