

Inducing novel endosymbioses by implanting bacteria in fungi
- PMID: 39358514
- PMCID: PMC11560845
- DOI: 10.1038/s41586-024-08010-x
Inducing novel endosymbioses by implanting bacteria in fungi
Abstract
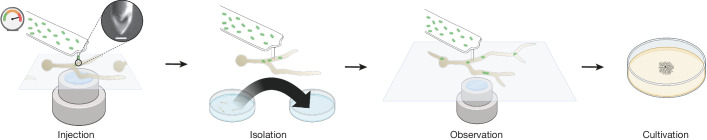
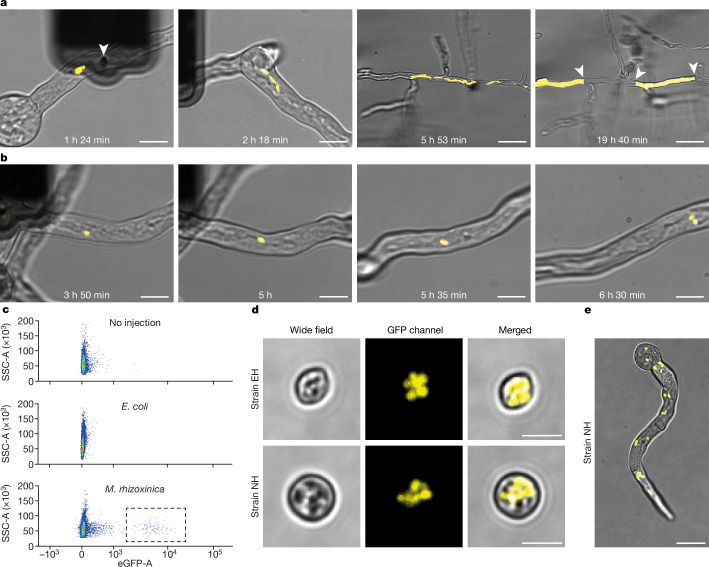
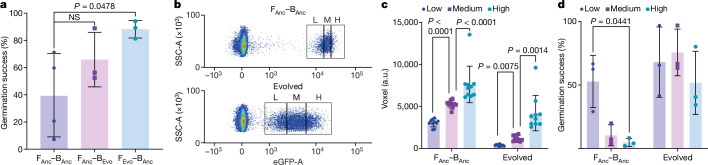
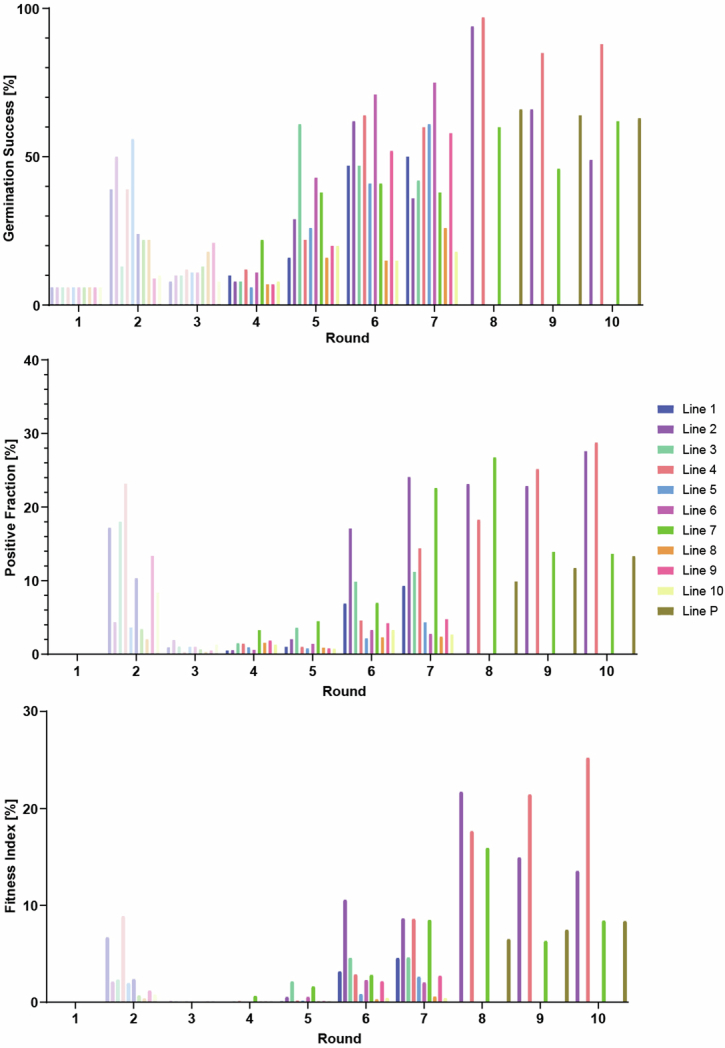
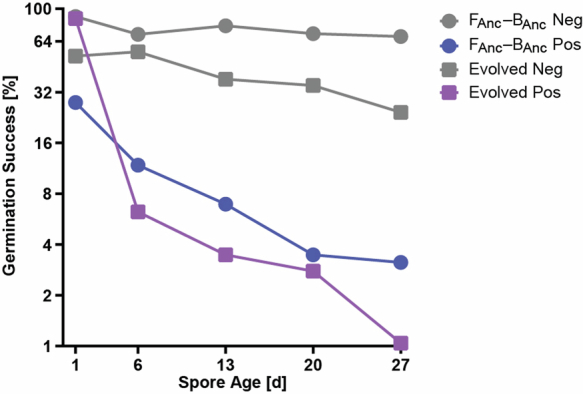
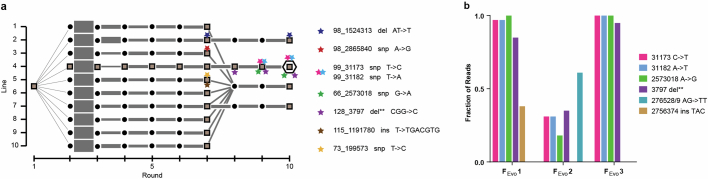
Endosymbioses have profoundly impacted the evolution of life and continue to shape the ecology of a wide range of species. They give rise to new combinations of biochemical capabilities that promote innovation and diversification1,2. Despite the many examples of known endosymbioses across the tree of life, their de novo emergence is rare and challenging to uncover in retrospect3-5. Here we implant bacteria into the filamentous fungus Rhizopus microsporus to follow the fate of artificially induced endosymbioses. Whereas Escherichia coli implanted into the cytosol induced septum formation, effectively halting endosymbiogenesis, Mycetohabitans rhizoxinica was transmitted vertically to the progeny at a low frequency. Continuous positive selection on endosymbiosis mitigated initial fitness constraints by several orders of magnitude upon adaptive evolution. Phenotypic changes were underscored by the accumulation of mutations in the host as the system stabilized. The bacterium produced rhizoxin congeners in its new host, demonstrating the transfer of a metabolic function through induced endosymbiosis. Single-cell implantation thus provides a powerful experimental approach to study critical events at the onset of endosymbiogenesis and opens opportunities for synthetic approaches towards designing endosymbioses with desired traits.
© 2024. The Author(s).
Conflict of interest statement
Figures



References
-
- Martin, W. & Müller, M. The hydrogen hypothesis for the first eukaryote. Nature392, 37–41 (1998). - PubMed
-
- Wernegreen, J. J. Endosymbiosis. Curr. Biol.22, 555–561 (2012). - PubMed
-
- Kobayashi, D. Y. & Crouch, J. A. Bacterial/fungal interactions: from pathogenes to mutualistic endosymbionts. Annu. Rev. Phytopathol.47, 63–82 (2009). - PubMed
-
- Meaney, R. S., Hamadache, S., Soltysiak, M. P. M. & Karas, B. J. Designer endosymbionts: converting free-living bacteria into organelles. Curr. Opin. Syst. Biol.24, 41–50 (2020).
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
