

Creation of de novo cryptic splicing for ALS and FTD precision medicine
- PMID: 39361759
- PMCID: PMC7616720
- DOI: 10.1126/science.adk2539
Creation of de novo cryptic splicing for ALS and FTD precision medicine
Abstract
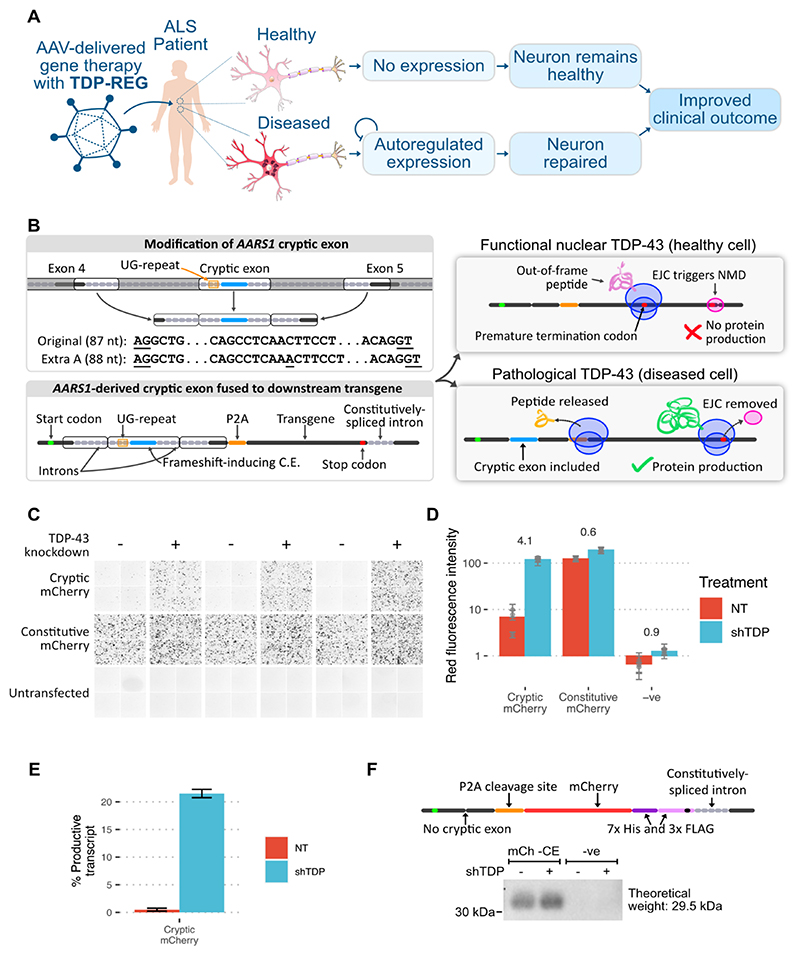
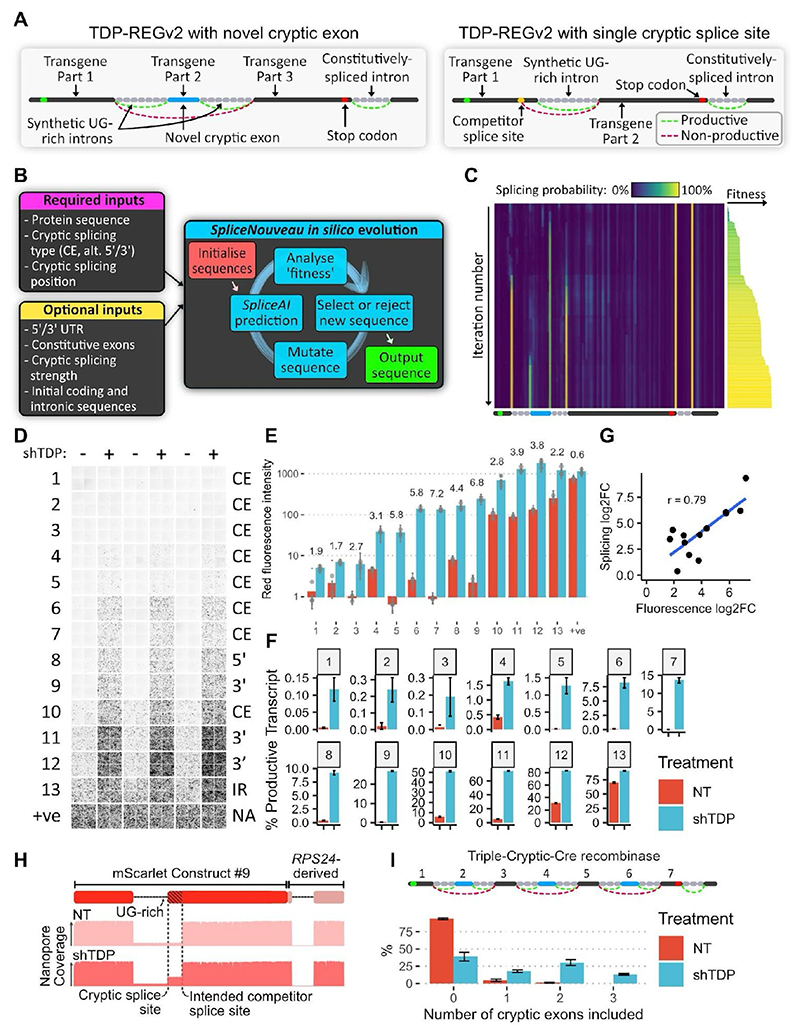
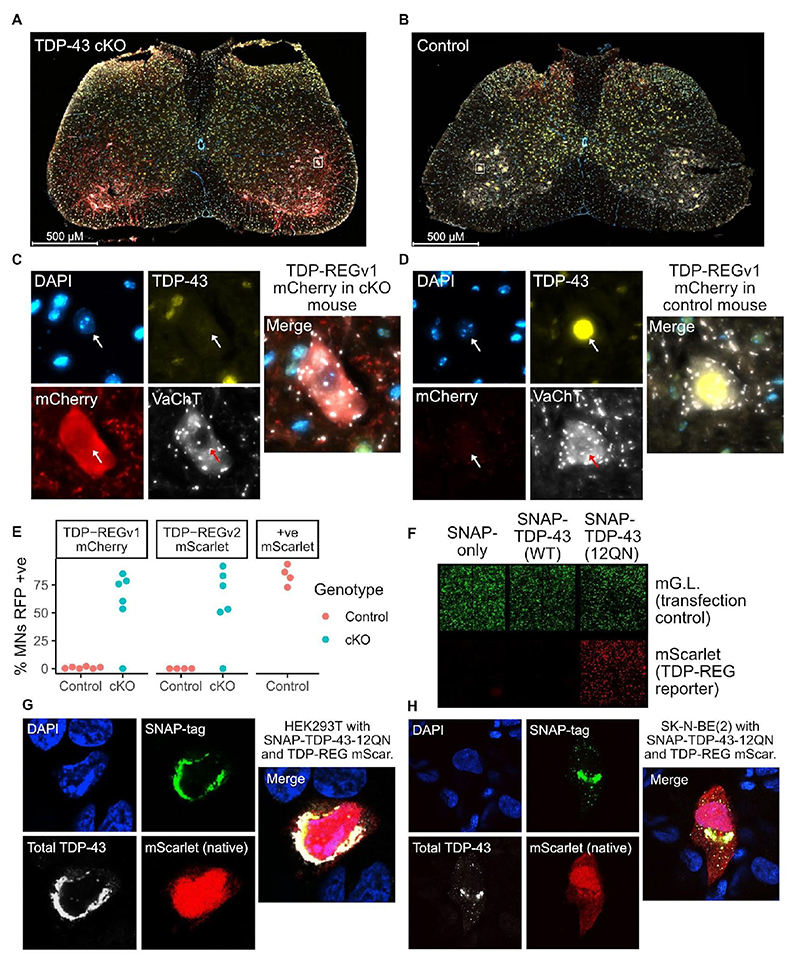
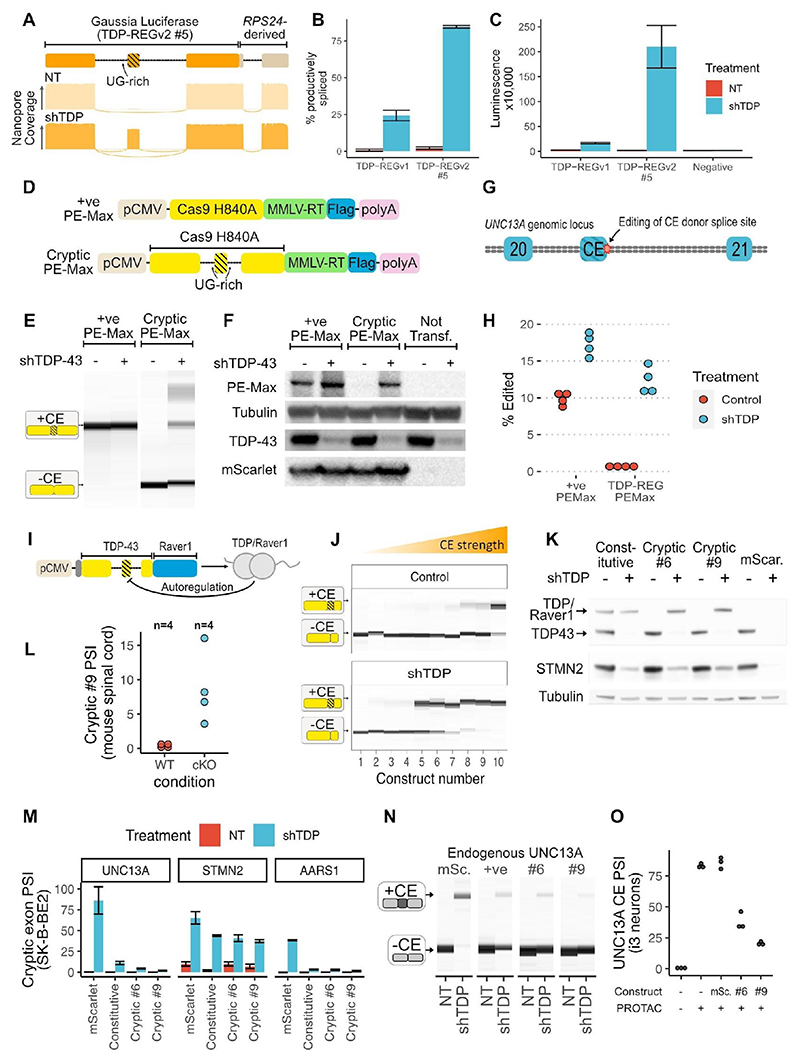
Loss of function of the RNA-binding protein TDP-43 (TDP-LOF) is a hallmark of amyotrophic lateral sclerosis (ALS) and other neurodegenerative disorders. Here we describe TDP-REG, which exploits the specificity of cryptic splicing induced by TDP-LOF to drive protein expression when and where the disease process occurs. The SpliceNouveau algorithm combines deep learning with rational design to generate customizable cryptic splicing events within protein-coding sequences. We demonstrate that expression of TDP-REG reporters is tightly coupled to TDP-LOF in vitro and in vivo. TDP-REG enables genomic prime editing to ablate the UNC13A cryptic donor splice site specifically upon TDP-LOF. Finally, we design TDP-REG vectors encoding a TDP-43/Raver1 fusion protein that rescues key pathological cryptic splicing events, paving the way for the development of precision therapies for TDP43-related disorders.
Conflict of interest statement
Figures
Update of
-
Creation of de novo cryptic splicing for ALS/FTD precision medicine.bioRxiv [Preprint]. 2023 Nov 15:2023.11.15.565967. doi: 10.1101/2023.11.15.565967. bioRxiv. 2023. Update in: Science. 2024 Oct 4;386(6717):61-69. doi: 10.1126/science.adk2539. PMID: 38014203 Free PMC article. Updated. Preprint.
Comment in
-
Regulators, mount up.Science. 2024 Oct 4;386(6717):24-25. doi: 10.1126/science.ads5951. Epub 2024 Oct 3. Science. 2024. PMID: 39361768
References
-
- Nelson PT, Dickson DW, Trojanowski JQ, Jack CR, Boyle PA, Arfanakis K, Rademakers R, Alafuzoff I, Attems J, Brayne C, Coyle-Gilchrist ITS, et al. Limbic-predominant age-related TDP-43 encephalopathy (LATE): consensus working group report. Brain. 2019;142:1503–1527. doi: 10.1093/brain/awz099. - DOI - PMC - PubMed
-
- Klim JR, Williams LA, Limone F, Guerra San Juan BN, Davis-Dusenbery BN, Mordes DA, Burberry A, Steinbaugh MJ, Gamage KK, Kirchner R, Moccia R, et al. ALS-implicated protein TDP-43 sustains levels of STMN2, a mediator of motor neuron growth and repair. Nat Neurosci. 2019;22:167–179. doi: 10.1038/s41593-018-0300-4. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
