

Bacterial RNA sensing by TLR8 requires RNase 6 processing and is inhibited by RNA 2'O-methylation
- PMID: 39363059
- PMCID: PMC11549399
- DOI: 10.1038/s44319-024-00281-9
Bacterial RNA sensing by TLR8 requires RNase 6 processing and is inhibited by RNA 2'O-methylation
Abstract
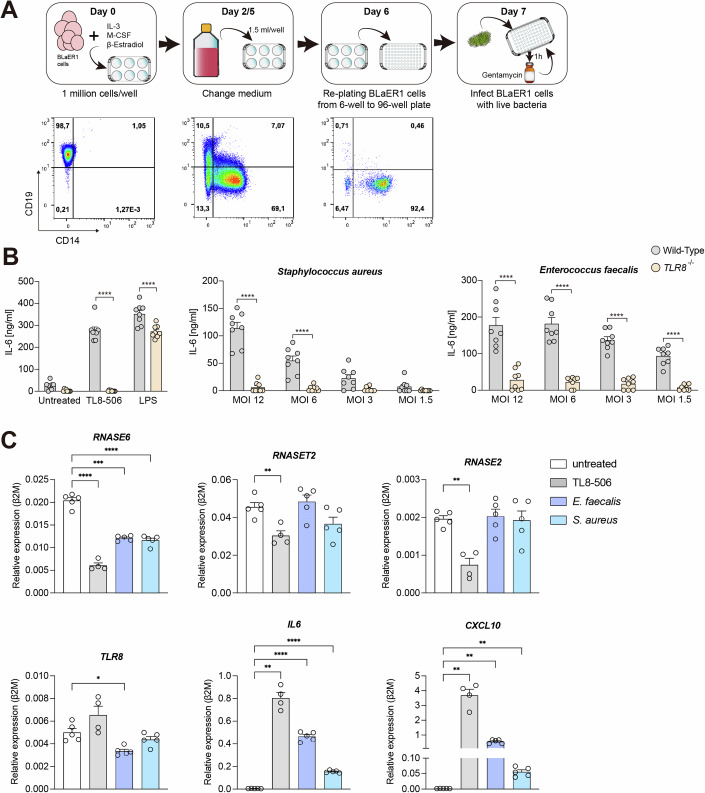
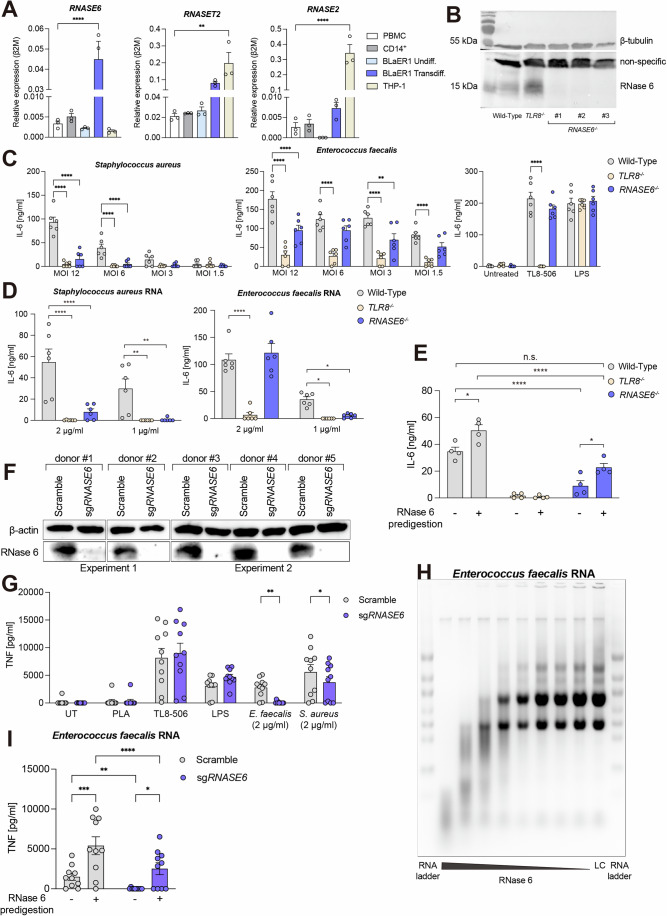
TLR8 senses single-stranded RNA (ssRNA) fragments, processed via cleavage by ribonuclease (RNase) T2 and RNase A family members. Processing by these RNases releases uridines and purine-terminated residues resulting in TLR8 activation. Monocytes show high expression of RNase 6, yet this RNase has not been analyzed for its physiological contribution to the recognition of bacterial RNA by TLR8. Here, we show a role for RNase 6 in TLR8 activation. BLaER1 cells, transdifferentiated into monocyte-like cells, as well as primary monocytes deficient for RNASE6 show a dampened TLR8-dependent response upon stimulation with isolated bacterial RNA (bRNA) and also upon infection with live bacteria. Pretreatment of bacterial RNA with recombinant RNase 6 generates fragments that induce TLR8 stimulation in RNase 6 knockout cells. 2'O-RNA methyl modification, when introduced at the first uridine in the UA dinucleotide, impairs processing by RNase 6 and dampens TLR8 stimulation. In summary, our data show that RNase 6 processes bacterial RNA and generates uridine-terminated breakdown products that activate TLR8.
Keywords: Innate Immunity; Nucleic Acid Recognition; RNA Modifications; RNase 6; Toll-Like Receptor 8.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Albanese M, Ruhle A, Mittermaier J, Mejías-Pérez E, Gapp M, Linder A, Schmacke NA, Hofmann K, Hennrich AA, Levy DN et al (2022) Rapid, efficient and activation-neutral gene editing of polyclonal primary human resting CD4+ T cells allows complex functional analyses. Nat Methods 19:81–89 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
