

Targeting CD38 with monoclonal antibodies disrupts key survival pathways in paediatric Burkitt's lymphoma malignant B cells
- PMID: 39364393
- PMCID: PMC11447455
- DOI: 10.1002/cti2.70011
Targeting CD38 with monoclonal antibodies disrupts key survival pathways in paediatric Burkitt's lymphoma malignant B cells
Abstract
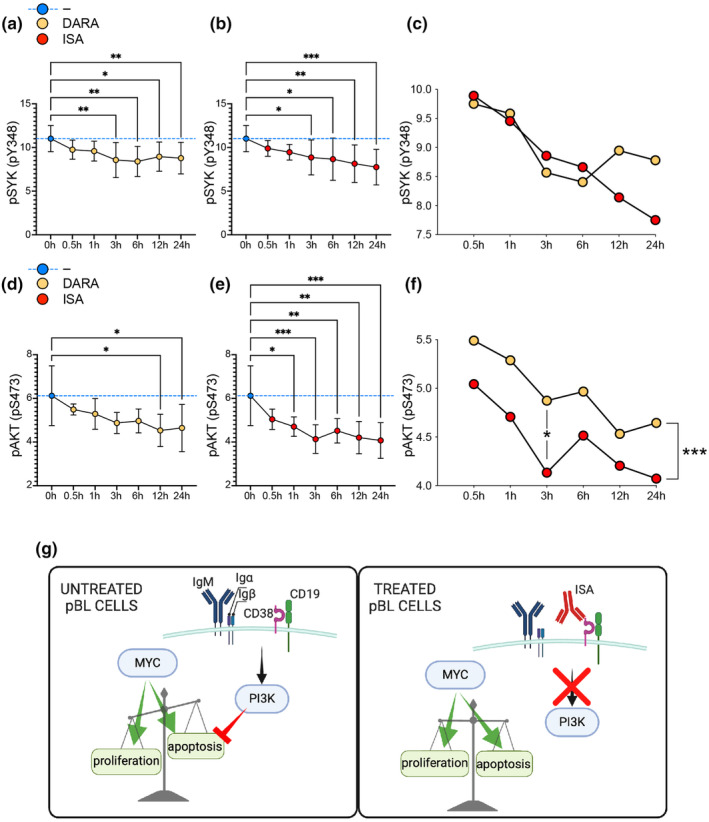
Objectives: Paediatric Burkitt's lymphoma (pBL) is the most common childhood non-Hodgkin B-cell lymphoma. Despite the encouraging survival rates for most children, treating cases with relapse/resistance to current therapies remains challenging. CD38 is a transmembrane protein highly expressed in pBL. This study investigates the effectiveness of CD38-targeting monoclonal antibodies (mAbs), daratumumab and isatuximab, in impairing crucial cellular processes and survival pathways in pBL malignant cells.
Methods: In silico analyses of patient samples, combined with in vitro experiments using the Ramos cell line, were conducted to assess the impact of daratumumab and isatuximab on cellular proliferation, apoptosis and the phosphoinositide 3-kinase (PI3K) pathway.
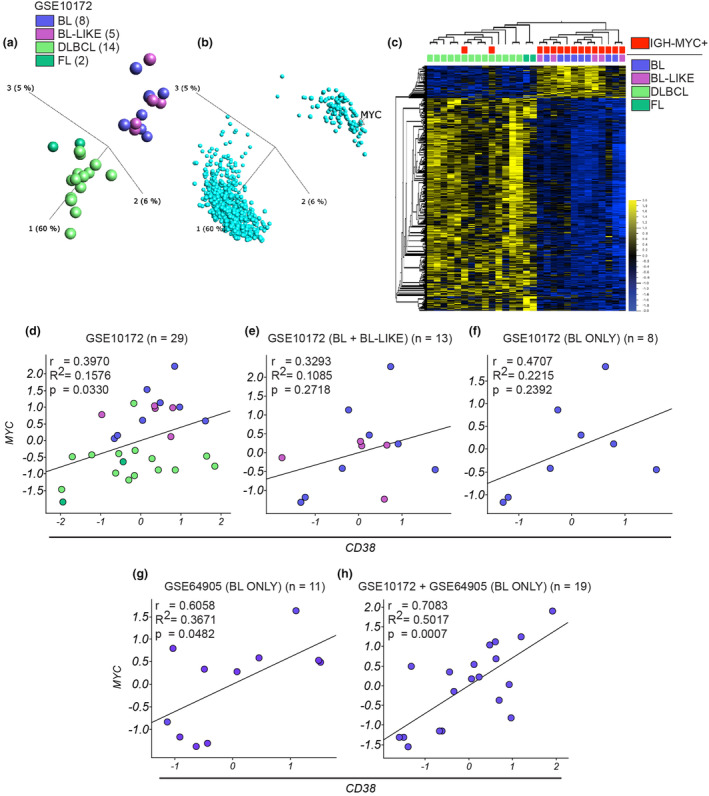
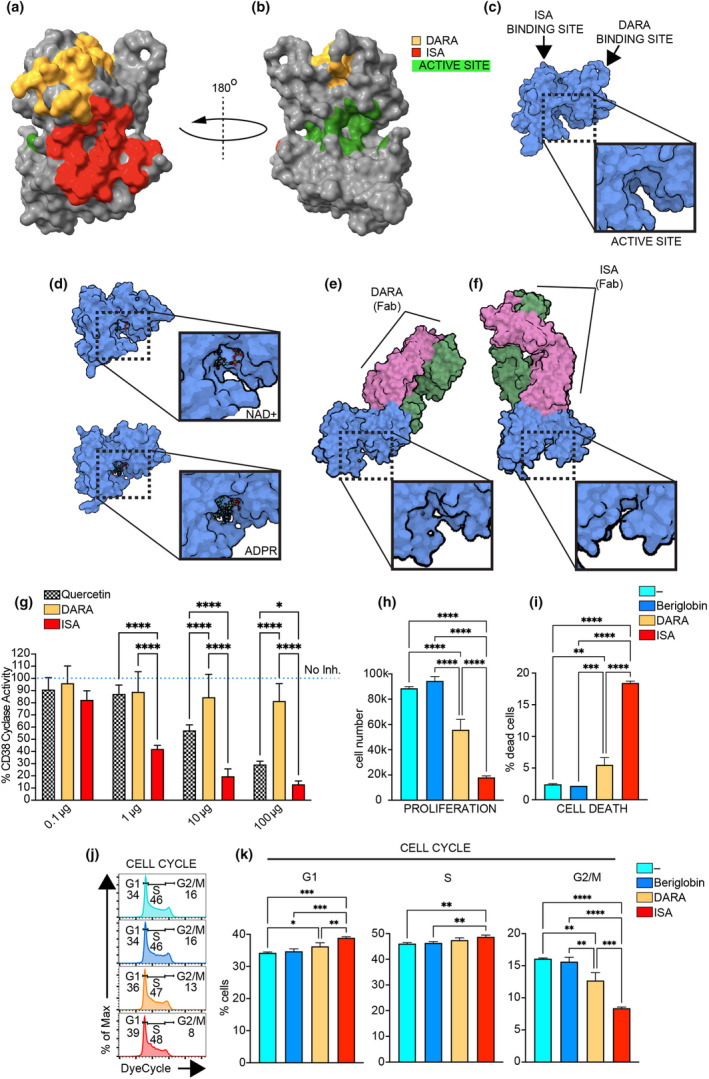
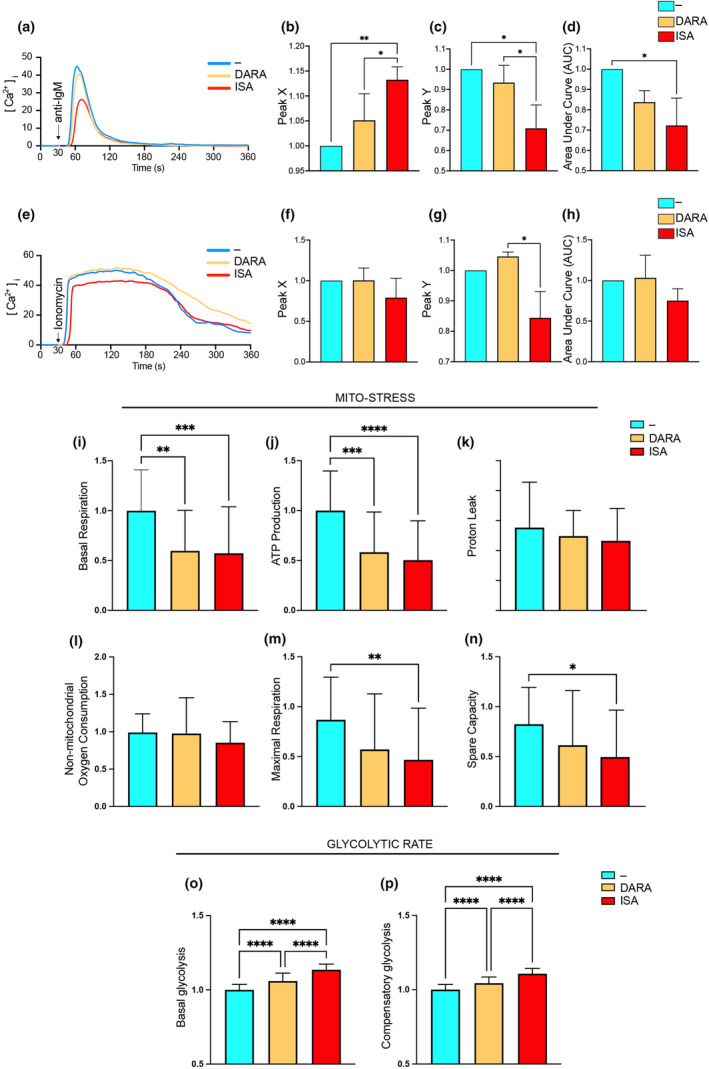
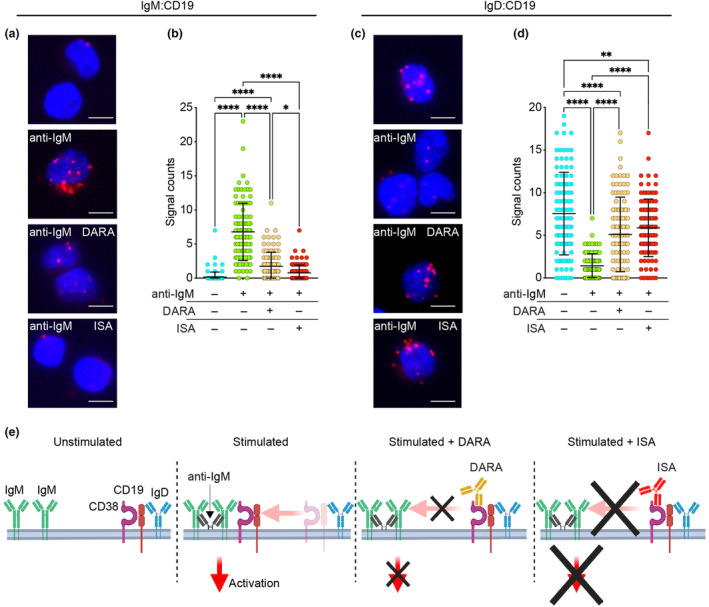
Results: Isatuximab was found to be more effective than daratumumab in disrupting B-cell receptor signalling, reducing cellular proliferation and inducing apoptosis. Additionally, isatuximab caused a significant impairment of the PI3K pathway and induced metabolic reprogramming in pBL cells. The study also revealed a correlation between CD38 and MYC expression levels in pBL patient samples, suggesting CD38 involvement in key oncogenic processes.
Conclusion: The study emphasises the therapeutic potential of CD38-targeting mAbs, particularly isatuximab, in pBL.
Keywords: CD38‐targeting monoclonal antibodies; chemotherapy resistance; daratumumab; immunotherapy; isatuximab; paediatric Burkitt's lymphoma.
© 2024 The Author(s). Clinical & Translational Immunology published by John Wiley & Sons Australia, Ltd on behalf of Australian and New Zealand Society for Immunology, Inc.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be considered as a potential conflict of interest. Cartoon figures were created with Biorender.com.
Figures
Similar articles
-
Anti-CD38 monoclonal antibody interference with blood compatibility testing: Differentiating isatuximab and daratumumab via functional epitope mapping.Transfusion. 2022 Nov;62(11):2334-2348. doi: 10.1111/trf.17137. Epub 2022 Oct 14. Transfusion. 2022. PMID: 36239134 Free PMC article.
-
Pharmacy considerations: Use of anti-CD38 monoclonal antibodies in relapsed and/or refractory multiple myeloma.J Oncol Pharm Pract. 2023 Jan;29(1):170-182. doi: 10.1177/10781552221107850. Epub 2022 Jun 20. J Oncol Pharm Pract. 2023. PMID: 35726199 Review.
-
FTL004, an anti-CD38 mAb with negligible RBC binding and enhanced pro-apoptotic activity, is a novel candidate for treatments of multiple myeloma and non-Hodgkin lymphoma.J Hematol Oncol. 2022 Dec 29;15(1):177. doi: 10.1186/s13045-022-01395-0. J Hematol Oncol. 2022. PMID: 36581954 Free PMC article.
-
CD46 and CD59 inhibitors enhance complement-dependent cytotoxicity of anti-CD38 monoclonal antibodies daratumumab and isatuximab in multiple myeloma and other B-cell malignancy cells.Cancer Biol Ther. 2024 Dec 31;25(1):2314322. doi: 10.1080/15384047.2024.2314322. Epub 2024 Feb 15. Cancer Biol Ther. 2024. PMID: 38361357 Free PMC article.
-
The Many Facets of CD38 in Lymphoma: From Tumor-Microenvironment Cell Interactions to Acquired Resistance to Immunotherapy.Cells. 2020 Mar 26;9(4):802. doi: 10.3390/cells9040802. Cells. 2020. PMID: 32225002 Free PMC article. Review.
References
-
- Gallamini A, Juweid M. Lymphoma [Internet]. Brisbane (AU): Exon Publications; 2021 Dec 28. PMID: 35226427. - PubMed
-
- López C, Burkhardt B, Chan JKC et al. Burkitt lymphoma. Nat Rev Dis Primers 2022; 8: 78. - PubMed
-
- Küppers R, Dalla‐Favera R. Mechanisms of chromosomal translocations in B cell lymphomas. Oncogene 2001; 20: 5580–5594. - PubMed
-
- Bras AE, Beishuizen A, Langerak AW et al. CD38 expression in paediatric leukaemia and lymphoma: Implications for antibody targeted therapy. Br J Haematol 2018; 180: 292–296. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials