

The role of structure in regulatory RNA elements
- PMID: 39364891
- PMCID: PMC11499389
- DOI: 10.1042/BSR20240139
The role of structure in regulatory RNA elements
Abstract
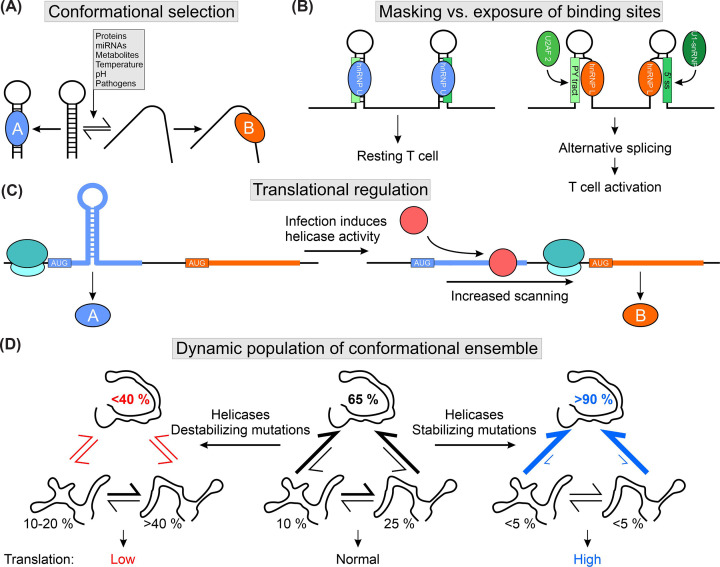
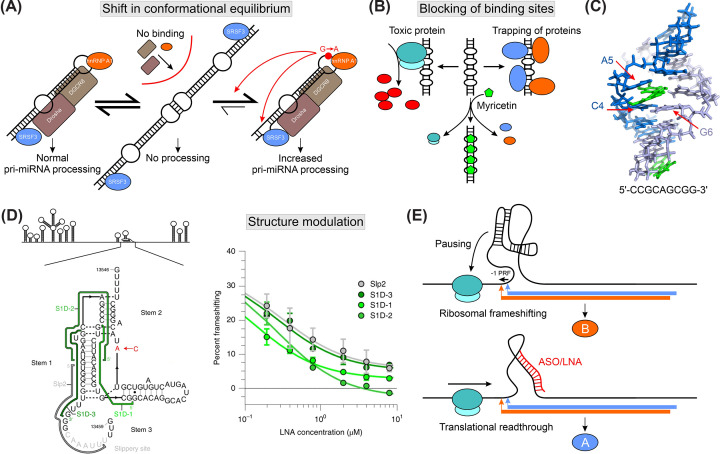
Regulatory RNA elements fulfill functions such as translational regulation, control of transcript levels, and regulation of viral genome replication. Trans-acting factors (i.e., RNA-binding proteins) bind the so-called cis elements and confer functionality to the complex. The specificity during protein-RNA complex (RNP) formation often exploits the structural plasticity of RNA. Functional integrity of cis-trans pairs depends on the availability of properly folded RNA elements, and RNA conformational transitions can cause diseases. Knowledge of RNA structure and the conformational space is needed for understanding complex formation and deducing functional effects. However, structure determination of RNAs under in vivo conditions remains challenging. This review provides an overview of structured eukaryotic and viral RNA cis elements and discusses the effect of RNA structural equilibria on RNP formation. We showcase implications of RNA structural changes for diseases, outline strategies for RNA structure-based drug targeting, and summarize the methodological toolbox for deciphering RNA structures.
Keywords: RNA conformers; RNA structure; RNA-binding proteins; cis-regulatory elements; dynamics; stem-loop.
© 2024 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures

Similar articles
-
NMR structures of loop B RNAs from the stem-loop IV domain of the enterovirus internal ribosome entry site: a single C to U substitution drastically changes the shape and flexibility of RNA.Biochemistry. 2004 May 18;43(19):5757-71. doi: 10.1021/bi0363228. Biochemistry. 2004. PMID: 15134450
-
KARR-seq reveals cellular higher-order RNA structures and RNA-RNA interactions.Nat Biotechnol. 2024 Dec;42(12):1909-1920. doi: 10.1038/s41587-023-02109-8. Epub 2024 Jan 18. Nat Biotechnol. 2024. PMID: 38238480 Free PMC article.
-
Nexus between RNA conformational dynamics and functional versatility.Curr Opin Struct Biol. 2024 Dec;89:102942. doi: 10.1016/j.sbi.2024.102942. Epub 2024 Oct 15. Curr Opin Struct Biol. 2024. PMID: 39413483 Review.
-
The structure and functions of coronavirus genomic 3' and 5' ends.Virus Res. 2015 Aug 3;206:120-33. doi: 10.1016/j.virusres.2015.02.025. Epub 2015 Feb 28. Virus Res. 2015. PMID: 25736566 Free PMC article. Review.
-
Identification and characterization of host factor interactions with cis-acting elements of rubella virus RNA.Arch Virol Suppl. 1994;9:255-67. doi: 10.1007/978-3-7091-9326-6_26. Arch Virol Suppl. 1994. PMID: 8032257
Cited by
-
5'-UTR G-Quadruplex-Mediated Translation Regulation in Eukaryotes: Current Understanding and Methodological Challenges.Int J Mol Sci. 2025 Jan 30;26(3):1187. doi: 10.3390/ijms26031187. Int J Mol Sci. 2025. PMID: 39940956 Free PMC article. Review.
-
Roquin exhibits opposing effects on RNA stem-loop stability through its two ROQ domain binding sites.Proc Natl Acad Sci U S A. 2025 Apr 15;122(15):e2424434122. doi: 10.1073/pnas.2424434122. Epub 2025 Apr 9. Proc Natl Acad Sci U S A. 2025. PMID: 40203046 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources