

The double-edged role of FASII regulator FabT in Streptococcus pyogenes infection
- PMID: 39366941
- PMCID: PMC11452403
- DOI: 10.1038/s41467-024-52637-3
The double-edged role of FASII regulator FabT in Streptococcus pyogenes infection
Abstract
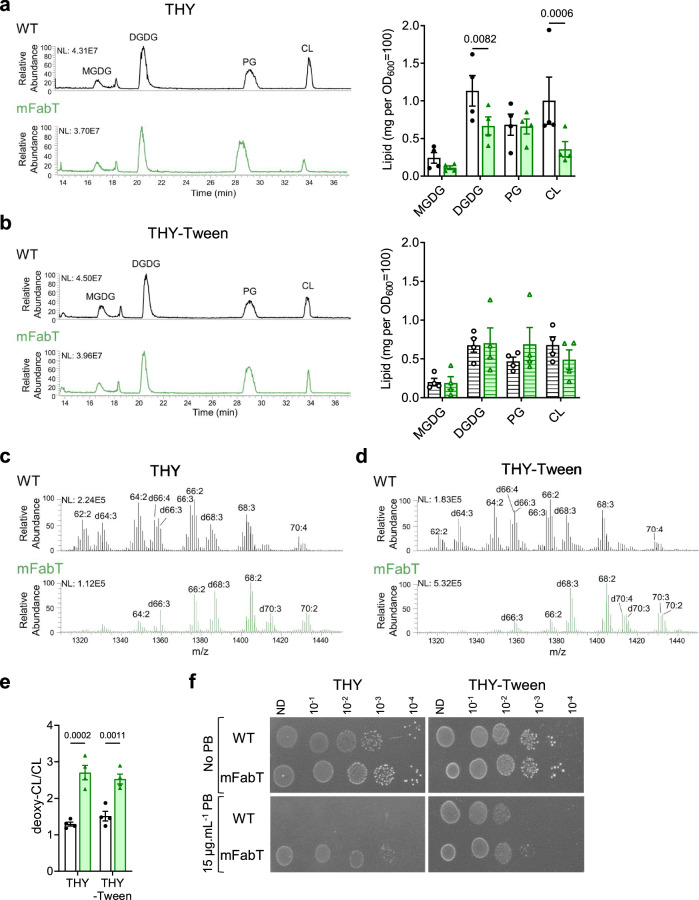
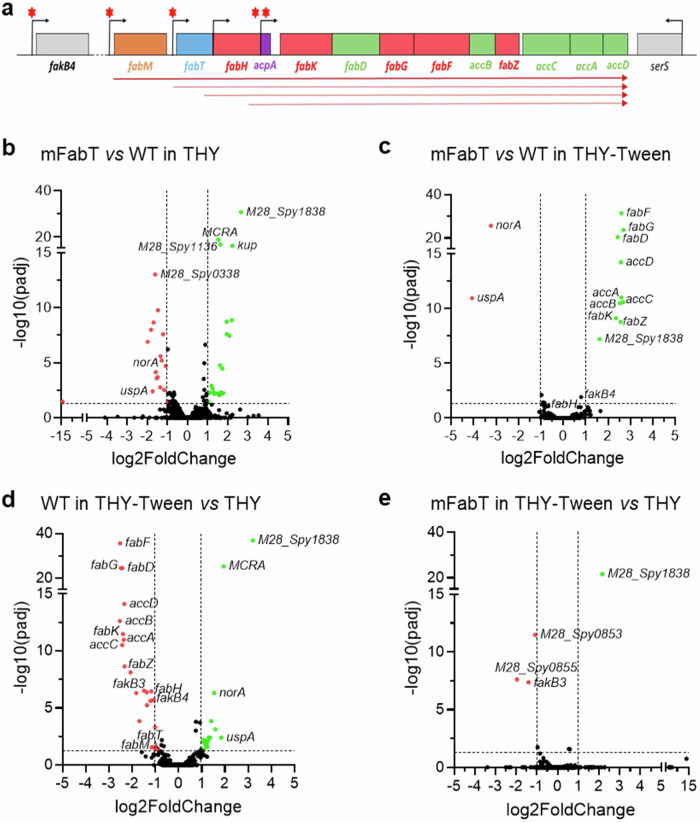
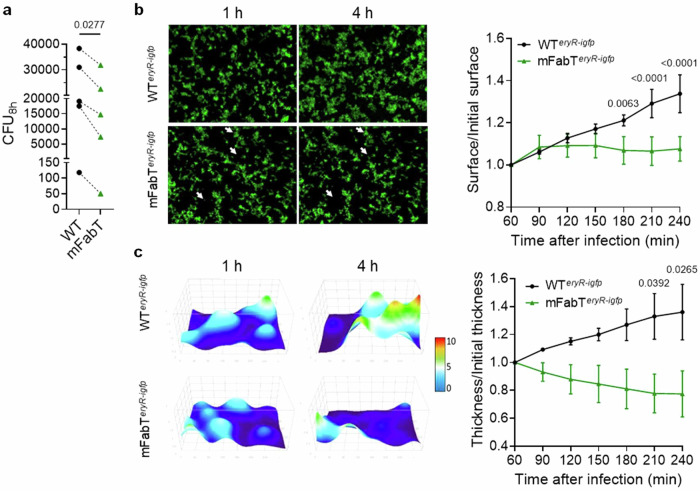
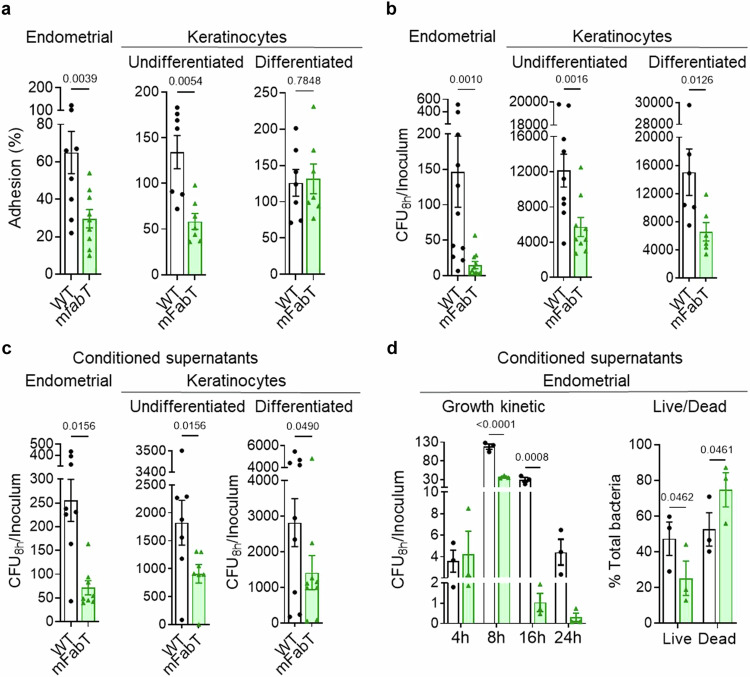
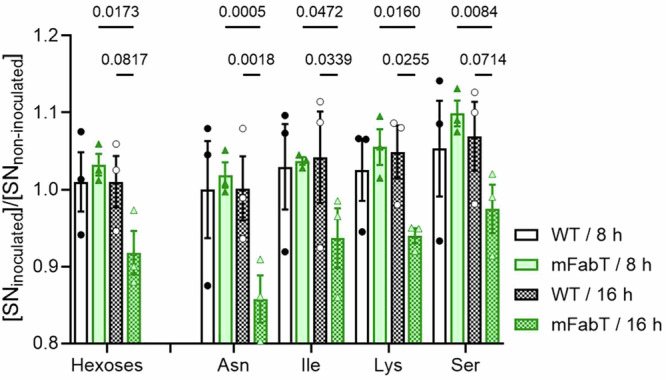
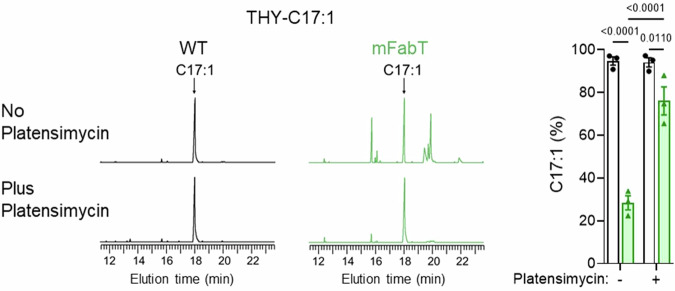
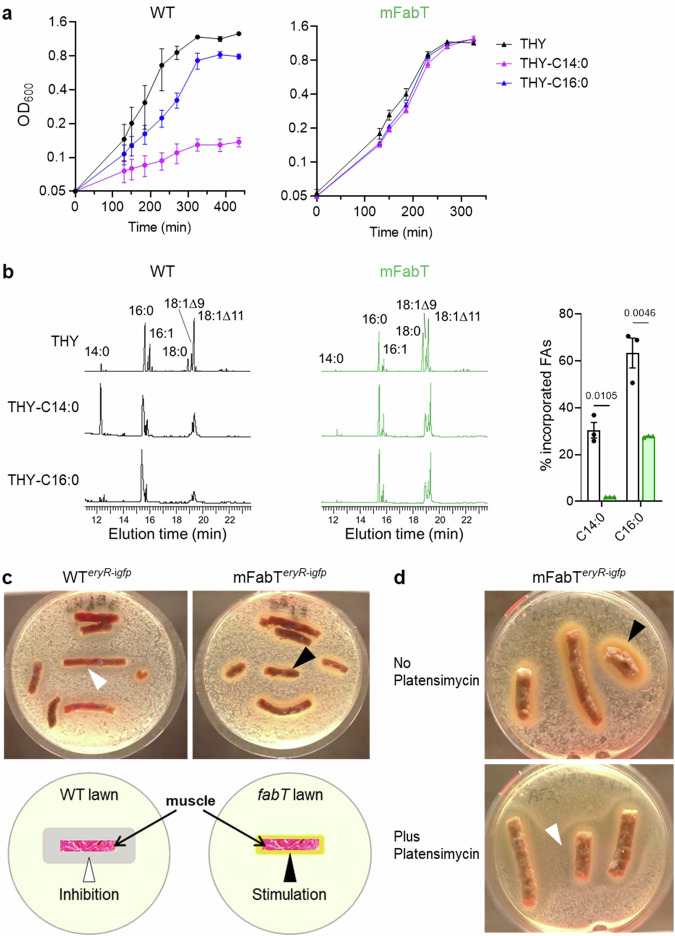
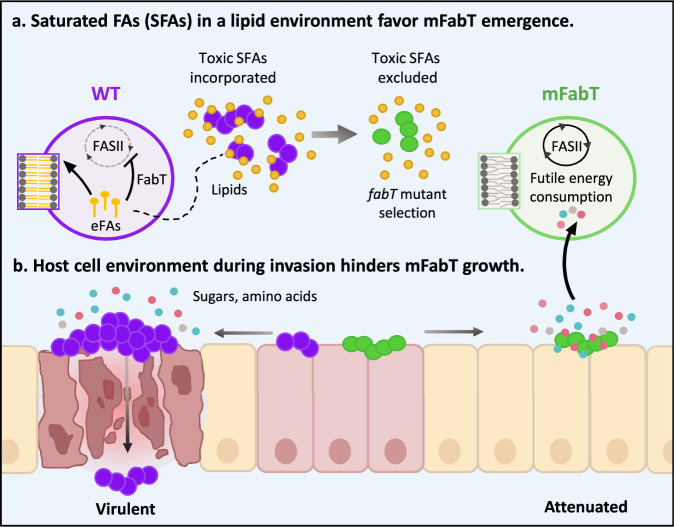
In Streptococcus pyogenes, the type II fatty acid (FA) synthesis pathway FASII is feedback-controlled by the FabT repressor bound to an acyl-Acyl carrier protein. Although FabT defects confer reduced virulence in animal models, spontaneous fabT mutants arise in vivo. We resolved this paradox by characterizing the conditions and mechanisms requiring FabT activity, and those promoting fabT mutant emergence. The fabT defect leads to energy dissipation, limiting mutant growth on human tissue products, which explains the FabT requirement during infection. Conversely, emerging fabT mutants show superior growth in biotopes rich in saturated FAs, where continued FASII activity limits their incorporation. We propose that membrane alterations and continued FASII synthesis are the primary causes for increased fabT mutant mortality in nutrient-limited biotopes, by failing to stop metabolic consumption. Our findings elucidate the rationale for emerging fabT mutants that improve bacterial survival in lipid-rich biotopes, but lead to a genetic impasse for infection.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- ANR-16-CE15-0013/Agence Nationale de la Recherche (French National Research Agency)
- ANR-22-AAMR-0007/Agence Nationale de la Recherche (French National Research Agency)
- DBF20161136769/Fondation pour la Recherche Médicale (Foundation for Medical Research in France)
- FDT202106012831/Fondation pour la Recherche Médicale (Foundation for Medical Research in France)
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
