

Concerted transcriptional regulation of the morphogenesis of hypothalamic neurons by ONECUT3
- PMID: 39366958
- PMCID: PMC11452682
- DOI: 10.1038/s41467-024-52762-z
Concerted transcriptional regulation of the morphogenesis of hypothalamic neurons by ONECUT3
Abstract
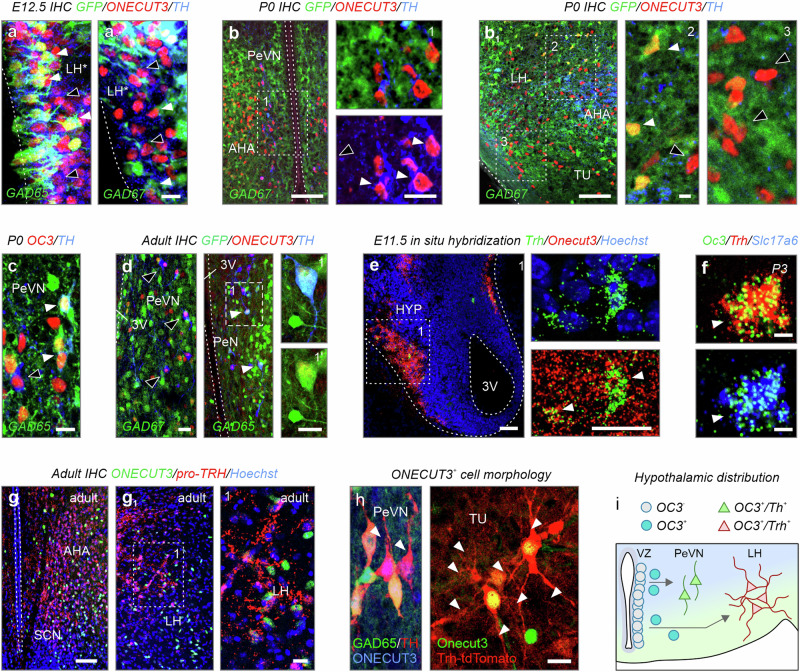
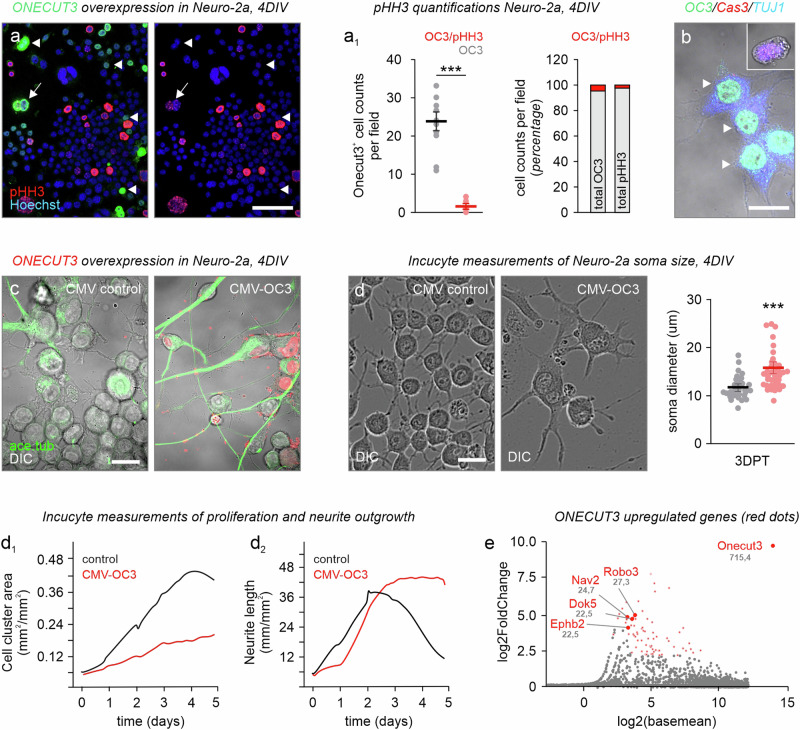
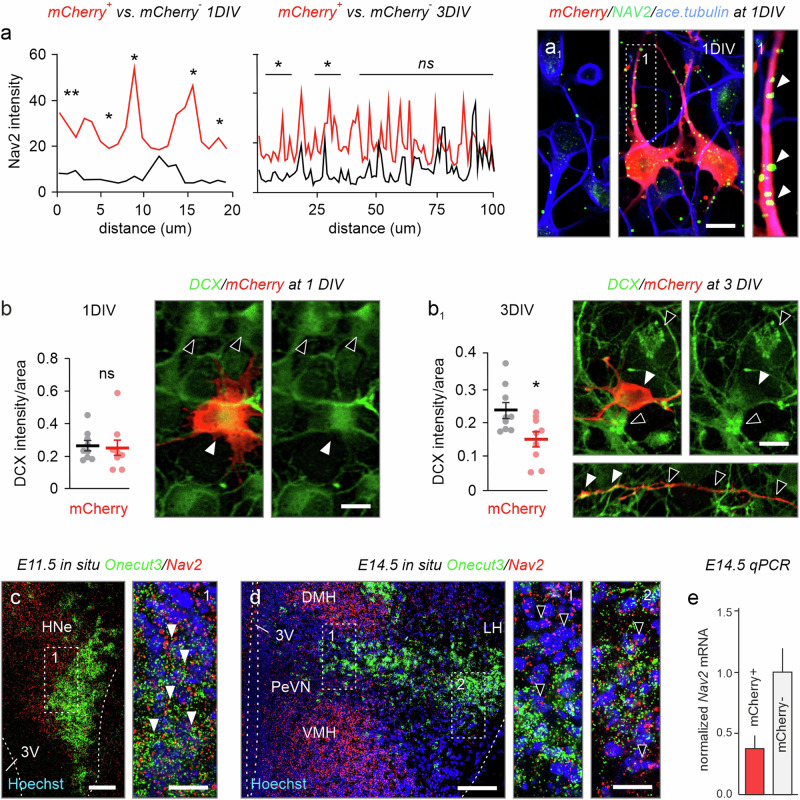
Acquisition of specialized cellular features is controlled by the ordered expression of transcription factors (TFs) along differentiation trajectories. Here, we find a member of the Onecut TF family, ONECUT3, expressed in postmitotic neurons that leave their Ascl1+/Onecut1/2+ proliferative domain in the vertebrate hypothalamus to instruct neuronal differentiation. We combined single-cell RNA-seq and gain-of-function experiments for gene network reconstruction to show that ONECUT3 affects the polarization and morphogenesis of both hypothalamic GABA-derived dopamine and thyrotropin-releasing hormone (TRH)+ glutamate neurons through neuron navigator-2 (NAV2). In vivo, siRNA-mediated knockdown of ONECUT3 in neonatal mice reduced NAV2 mRNA, as well as neurite complexity in Onecut3-containing neurons, while genetic deletion of Onecut3/ceh-48 in C. elegans impaired neurocircuit wiring, and sensory discrimination-based behaviors. Thus, ONECUT3, conserved across neuronal subtypes and many species, underpins the polarization and morphological plasticity of phenotypically distinct neurons that descend from a common pool of Ascl1+ progenitors in the hypothalamus.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
Brain-wide mapping of efferent projections of glutamatergic (Onecut3+ ) neurons in the lateral mouse hypothalamus.Acta Physiol (Oxf). 2023 Jul;238(3):e13973. doi: 10.1111/apha.13973. Epub 2023 Apr 25. Acta Physiol (Oxf). 2023. PMID: 37029761 Free PMC article.
-
A transcription factor network specifying inhibitory versus excitatory neurons in the dorsal spinal cord.Development. 2014 Jul;141(14):2803-12. doi: 10.1242/dev.105866. Epub 2014 Jun 12. Development. 2014. PMID: 24924197 Free PMC article.
-
Induction of specific neuron types by overexpression of single transcription factors.In Vitro Cell Dev Biol Anim. 2016 Oct;52(9):961-973. doi: 10.1007/s11626-016-0056-7. Epub 2016 Jun 1. In Vitro Cell Dev Biol Anim. 2016. PMID: 27251161 Free PMC article.
-
Ascl1 is required to specify a subset of ventromedial hypothalamic neurons.Development. 2020 May 28;147(10):dev180067. doi: 10.1242/dev.180067. Development. 2020. PMID: 32253239
-
Making neurons, made easy: The use of Neurogenin-2 in neuronal differentiation.Stem Cell Reports. 2022 Jan 11;17(1):14-34. doi: 10.1016/j.stemcr.2021.11.015. Epub 2021 Dec 30. Stem Cell Reports. 2022. PMID: 34971564 Free PMC article. Review.
Cited by
-
Molecular Fingerprint of Endocannabinoid Signaling in the Developing Paraventricular Nucleus of the Hypothalamus as Revealed by Single-Cell RNA-Seq and In Situ Hybridization.Cells. 2025 May 27;14(11):788. doi: 10.3390/cells14110788. Cells. 2025. PMID: 40497964 Free PMC article.
References
-
- Sower, S. A., Freamat, M. & Kavanaugh, S. I. The origins of the vertebrate hypothalamic-pituitary-gonadal (HPG) and hypothalamic-pituitary-thyroid (HPT) endocrine systems: new insights from lampreys. Gen. Comp. Endocrinol.161, 20–29 (2009). - PubMed
-
- van den Pol, A. N., Obrietan, K., Cao, V. & Trombley, P. Q. Embryonic hypothalamic expression of functional glutamate receptors. Neuroscience67, 419–439 (1995). - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- 2018-02838/Vetenskapsrådet (Swedish Research Council)
- NNF20OC0053667/Novo Nordisk Fonden (Novo Nordisk Foundation)
- ERC-2020-AdG-101021016/EC | EC Seventh Framework Programm | FP7 Ideas: European Research Council (FP7-IDEAS-ERC - Specific Programme: "Ideas" Implementing the Seventh Framework Programme of the European Community for Research, Technological Development and Demonstration Activities (2007 to 2013))
- P 34121-B/Austrian Science Fund (Fonds zur Förderung der Wissenschaftlichen Forschung)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
