

Peroxisomal homeostasis in metabolic diseases and its implication in ferroptosis
- PMID: 39367496
- PMCID: PMC11451054
- DOI: 10.1186/s12964-024-01862-w
Peroxisomal homeostasis in metabolic diseases and its implication in ferroptosis
Abstract
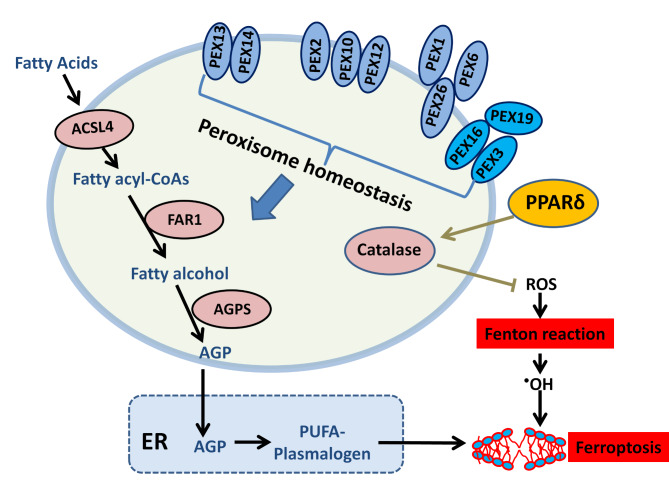
Peroxisomes are dynamic organelles involved in various cellular processes, including lipid metabolism, redox homeostasis, and intracellular metabolite transfer. Accumulating evidence suggests that peroxisomal homeostasis plays a crucial role in human health and disease, particularly in metabolic disorders and ferroptosis. The abundance and function of peroxisomes are regulated by a complex interplay between biogenesis and degradation pathways, involving peroxins, membrane proteins, and pexophagy. Peroxisome-dependent lipid metabolism, especially the synthesis of ether-linked phospholipids, has been implicated in modulating cellular susceptibility to ferroptosis, a newly discovered form of iron-dependent cell death. This review discusses the current understanding of peroxisome homeostasis, its roles in redox regulation and lipid metabolism, and its implications in human diseases. We also summarize the main mechanisms of ferroptosis and highlight recent discoveries on how peroxisome-dependent metabolism and signaling influence ferroptosis sensitivity. A better understanding of the interplay between peroxisomal homeostasis and ferroptosis may provide new insights into disease pathogenesis and reveal novel therapeutic strategies for peroxisome-related metabolic disorders and ferroptosis-associated diseases.
Keywords: Ferroptosis; Homeostasis; Lipid metabolism; Metabolic disorders; Peroxisome; Redox regulation; Therapeutic strategies.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
