

Docosahexaenoic acid (DHA) is a driving force regulating gene expression in bluefin tuna (Thunnus thynnus) larvae development
- PMID: 39369082
- PMCID: PMC11455926
- DOI: 10.1038/s41598-024-74152-7
Docosahexaenoic acid (DHA) is a driving force regulating gene expression in bluefin tuna (Thunnus thynnus) larvae development
Erratum in
-
Author Correction: Docosahexaenoic acid (DHA) is a driving force regulating gene expression in bluefin tuna (Thunnus thynnus) larvae development.Sci Rep. 2024 Dec 2;14(1):29970. doi: 10.1038/s41598-024-80623-8. Sci Rep. 2024. PMID: 39622861 Free PMC article. No abstract available.
Abstract
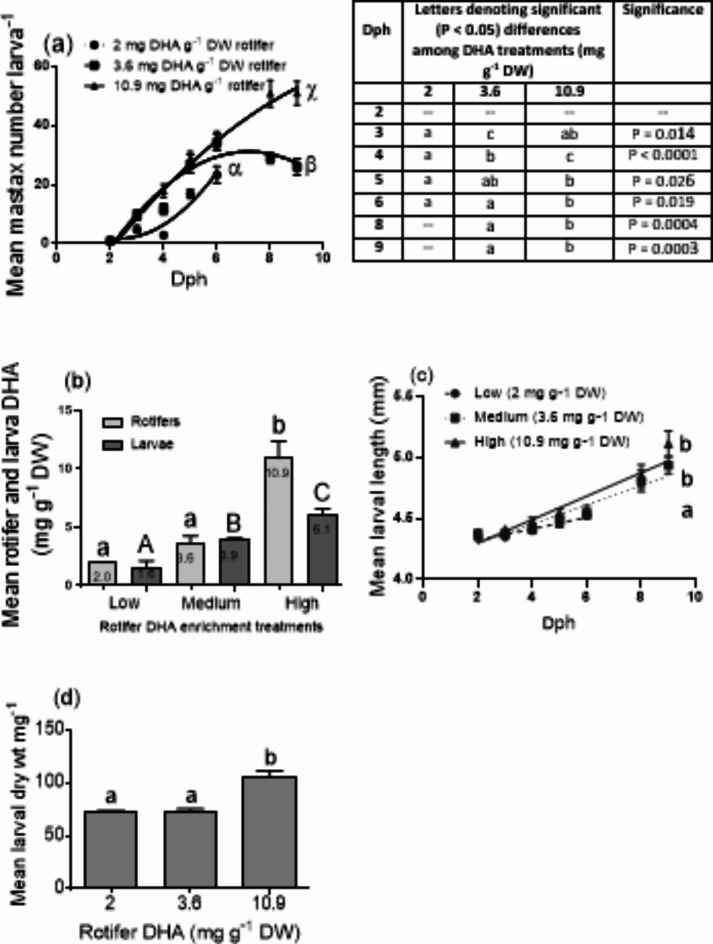
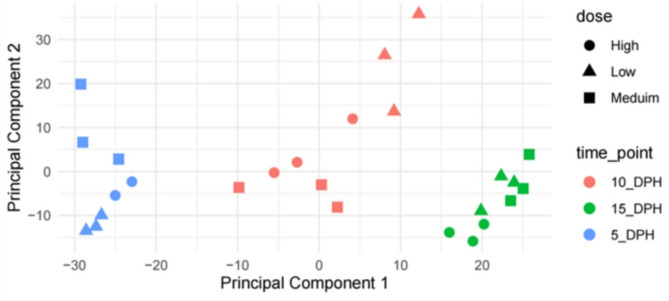
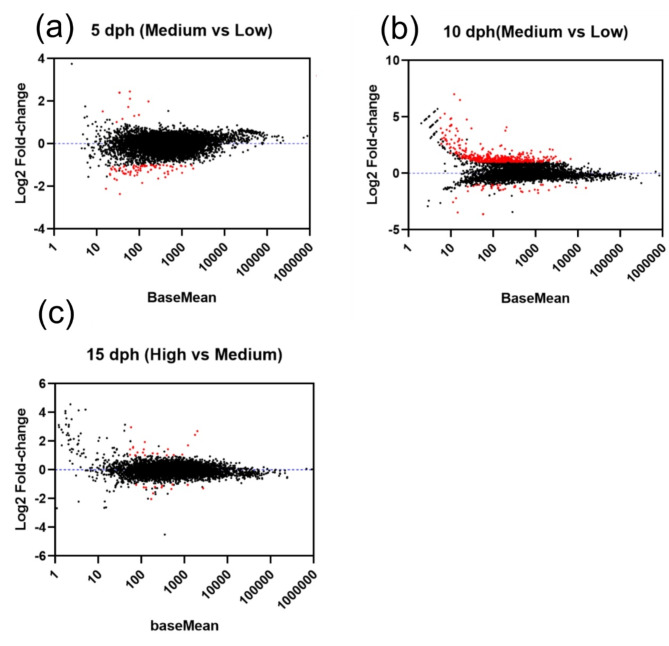
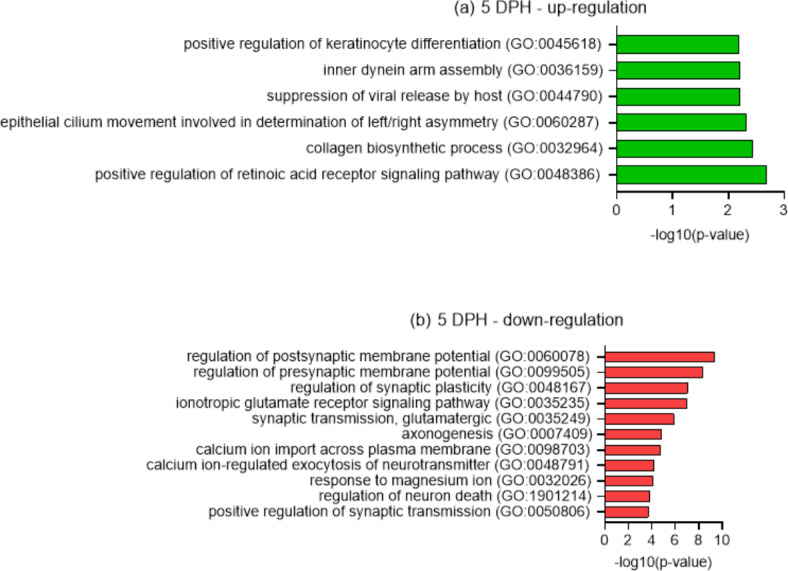
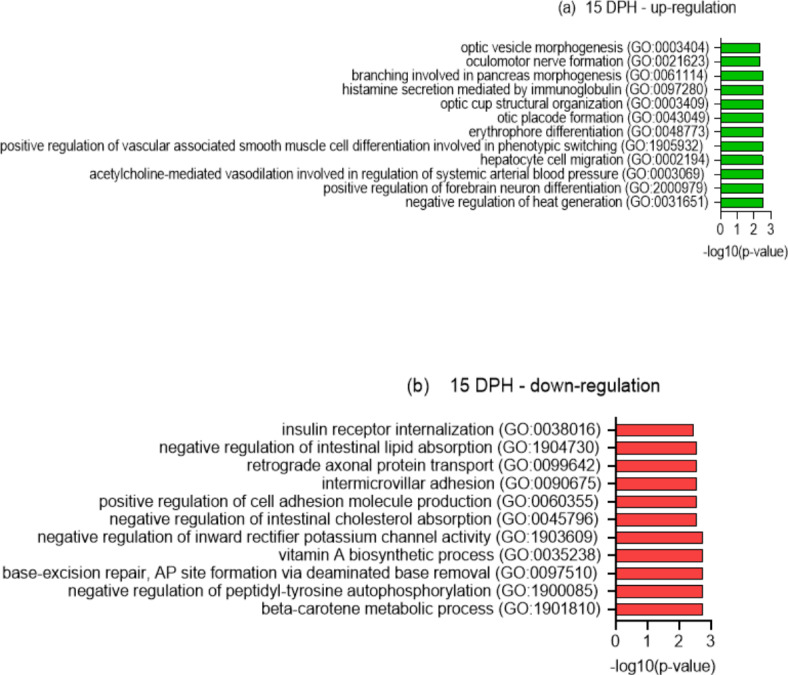
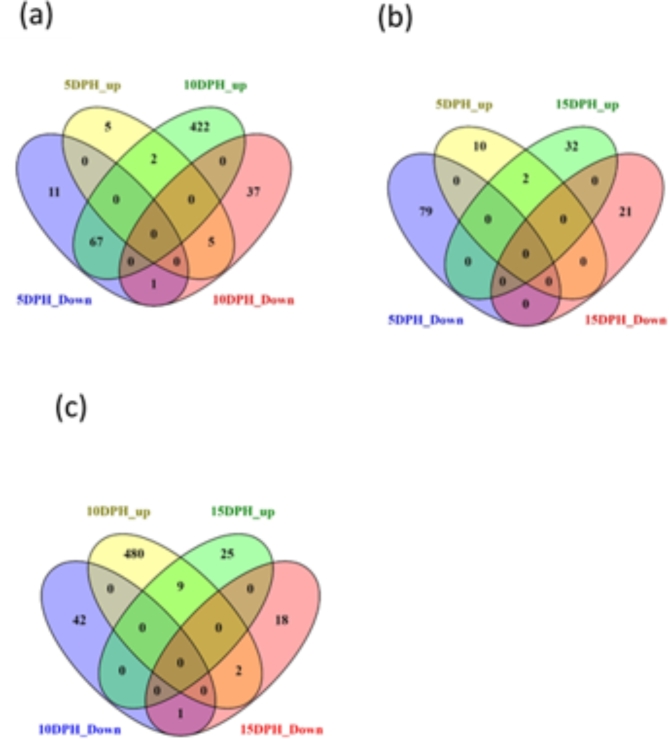
This study elucidated the role of DHA-modulated genes in the development and growth of Atlantic bluefin tuna (Thunnus thynnus) larvae ingesting increasing levels of DHA in their rotifer prey. The effect of feeding low, medium, and high rotifer (Brachionus rotundiformis) DHA levels (2.0, 3.6 and 10.9 mg DHA g-1 DW, respectively) was tested on 2-15 days post hatching (dph) bluefin tuna larvae. Larval DHA content markedly (P < 0.05) increased in a DHA dose-dependent manner (1.5, 3.9, 6.1 mg DHA g-1 DW larva, respectively), that was positively correlated with larval prey consumption and growth (P < 0.05). Gene ontology enrichment analyses of differentially expressed genes (DEGs) demonstrated dietary DHA significantly (P < 0.05) affected different genes and biological processes at different developmental ages. The number of DHA up-regulated DEGs was highest in 10 dph larvae (491), compared to 5 (12) and 15 dph fish (34), and were mainly involved in neural and synaptic development in the brain and spinal cord. In contrast, DHA in older 15 dph larvae elicited fewer DEGs but played critical roles over a wider range of developing organs. The emerging picture underscores the importance of DHA-modulated gene expression as a driving force in bluefin tuna larval development and growth.
Keywords: Bluefin tuna; Docosahexaenoic acid; Gene regulation; Larvae; Neurons; Synaptic function.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Izquierdo, M. & Koven, W. Lipids. In Larval fish nutrition 47–81 (Wiley, 2011). 10.1002/9780470959862.ch2.
-
- Gaon, A. et al. The combined DHA and taurine effect on vision, prey capture and growth in different age larvae of gilthead sea bream (Sparus aurata). Aquaculture545, 737181 (2021). - DOI
-
- Bell, M. V. et al. Dietary deficiency of docosahexaenoic acid impairs vision at low light intensities in juvenile herring (Clupea harengus L.). Lipids30, (1995). - PubMed
-
- Benítez-Santana, T. et al. Dietary n-3 HUFA deficiency induces a reduced visual response in gilthead seabream Sparus aurata larvae. Aquaculture264, 408–417 (2007). - DOI
-
- Mourente, G. et al. Lipids in female northern bluefin tuna (Thunnus thynnus thynnus L.) during sexual maturation. Fish Physiol. Biochem.2, 351–363 (2002).
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
