

EZH2 functional dichotomy in reactive oxygen species-stratified glioblastoma
- PMID: 39373211
- PMCID: PMC11812038
- DOI: 10.1093/neuonc/noae206
EZH2 functional dichotomy in reactive oxygen species-stratified glioblastoma
Abstract
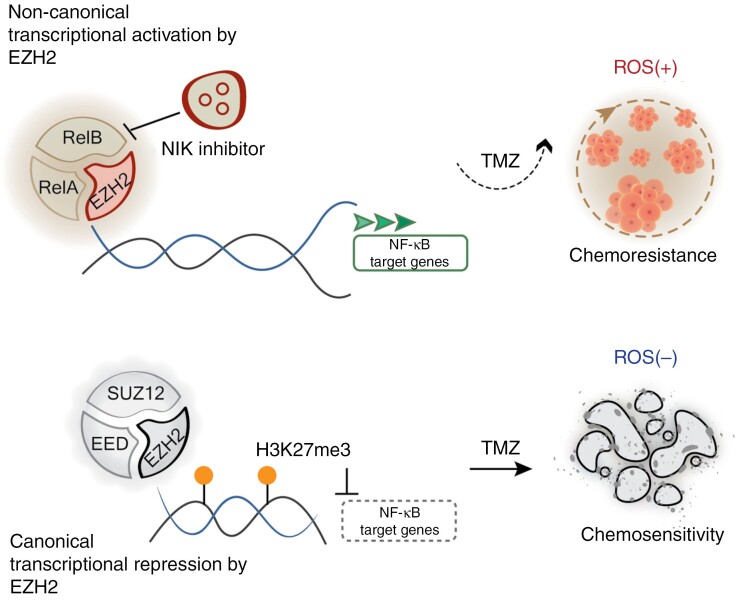
Background: Enhancer of zeste homolog 2 (EZH2), well known for its canonical methyltransferase activity in transcriptional repression in many cancers including glioblastoma (GBM), has an understudied noncanonical function critical for sustained tumor growth. Recent GBM consortial efforts reveal complex molecular heterogeneity for which therapeutic vulnerabilities correlated with subtype stratification remain relatively unexplored. Current enzymatic EZH2 inhibitors (EZH2inh) targeting its canonical su(var)3-9, enhancer-of-zeste and trithorax domain show limited efficacy and lack durable response, suggesting that underlying differences in the noncanonical pathway may yield new knowledge. Here, we unveiled dual roles of the EZH2 CXC domain in therapeutically distinct, reactive oxygen species (ROS)-stratified tumors.
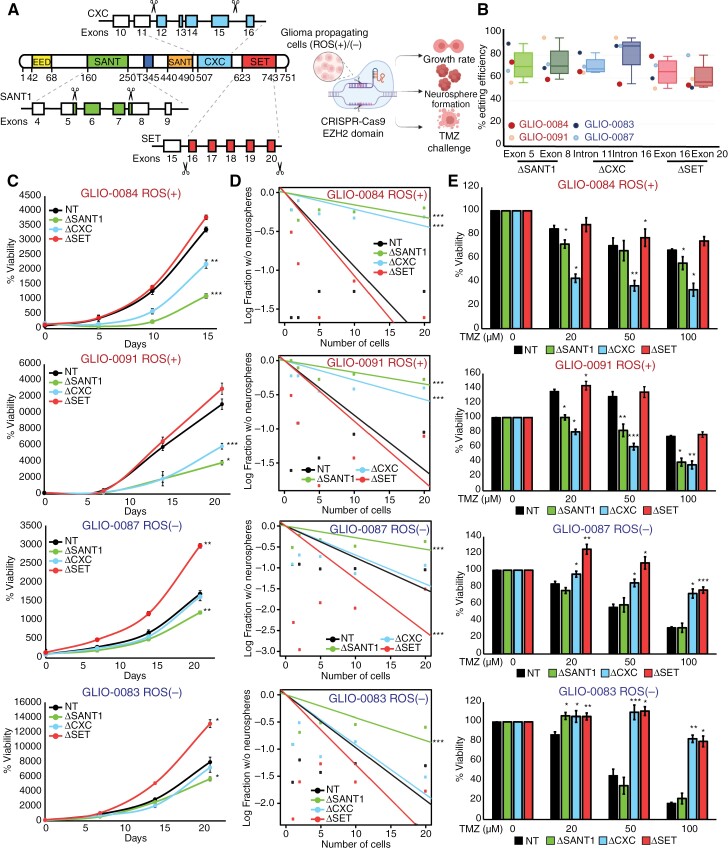
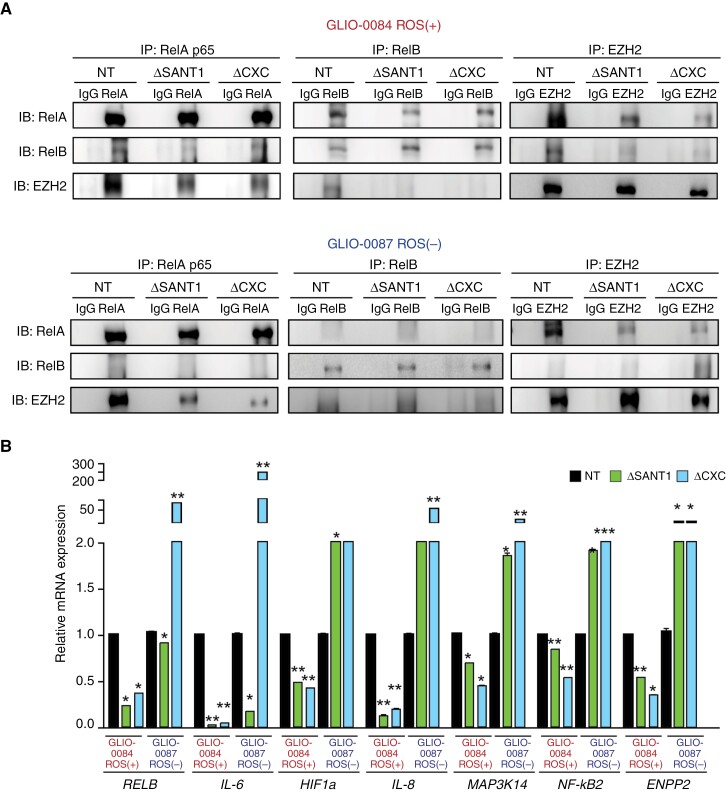
Methods: We analyzed differentially expressed genes between ROS classes by examining cis-regulatory elements as well as clustering of activities and pathways to identify EZH2 as the key mediator in ROS-stratified cohorts. Pull-down assays and CRISPR knockout of EZH2 domains were used to dissect the distinct functions of EZH2 in ROS-stratified GBM cells. The efficacy of NF-κB-inducing kinase inhibitor (NIKinh) and standard-of-care temozolomide was evaluated using orthotopic patient-derived GBM xenografts.
Results: In ROS(+) tumors, CXC-mediated co-interaction with RelB drives constitutive activation of noncanonical NF-κB2 signaling, sustaining the ROS(+) chemoresistant phenotype. In contrast, in ROS(-) subtypes, Polycomb Repressive Complex 2 methyltransferase activity represses canonical NF-κB. Addressing the lack of EZH2inh targeting its nonmethyltransferase roles, we utilized a brain-penetrant NIKinh that disrupts EZH2-RelB binding, consequently prolonging survival in orthotopic ROS(+)-implanted mice.
Conclusions: Our findings highlight the functional dichotomy of the EZH2 CXC domain in governing ROS-stratified therapeutic resistance, thereby advocating for the development of therapeutic approaches targeting its noncanonical activities and underscoring the significance of patient stratification methodologies.
Keywords: EZH2; brain cancer; glioblastoma; patient stratification; precision medicine.
© The Author(s) 2024. Published by Oxford University Press on behalf of the Society for Neuro-Oncology.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
