

Amalgam plays a dual role in controlling the number of leg muscle progenitors and regulating their interactions with the developing Drosophila tendon
- PMID: 39374263
- PMCID: PMC11486429
- DOI: 10.1371/journal.pbio.3002842
Amalgam plays a dual role in controlling the number of leg muscle progenitors and regulating their interactions with the developing Drosophila tendon
Abstract
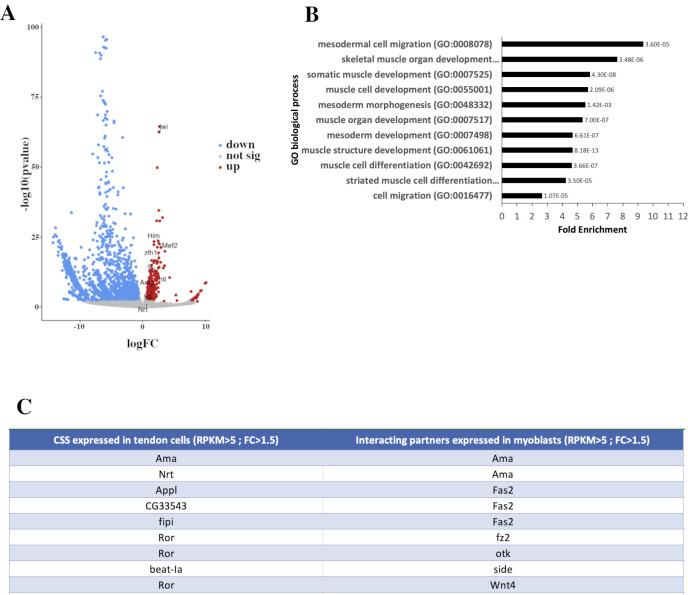
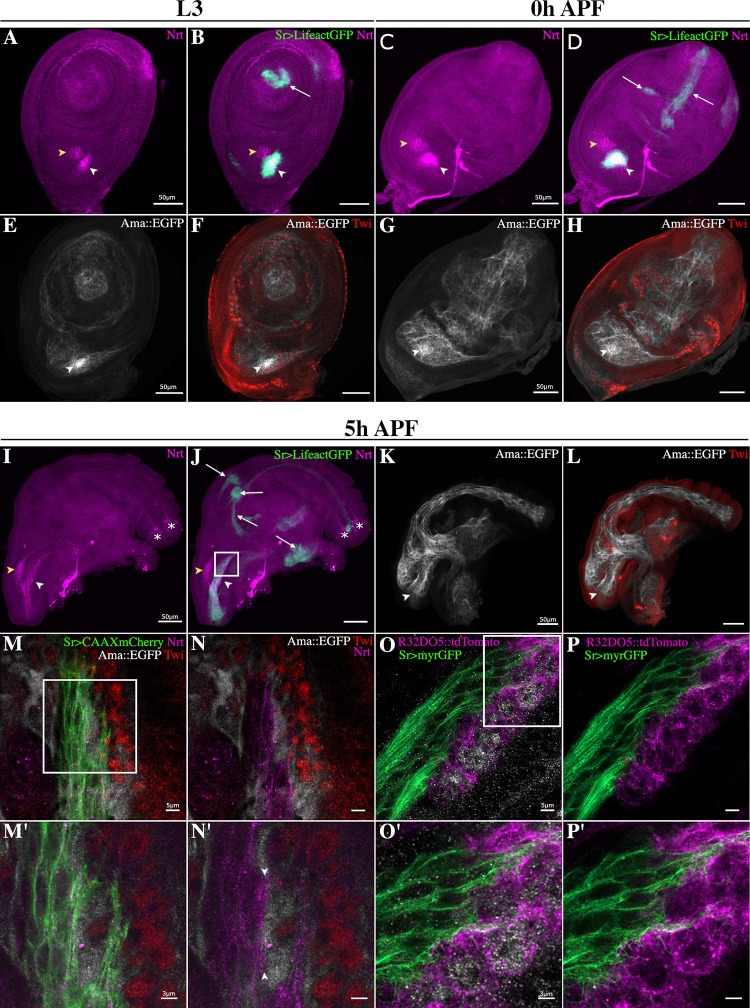
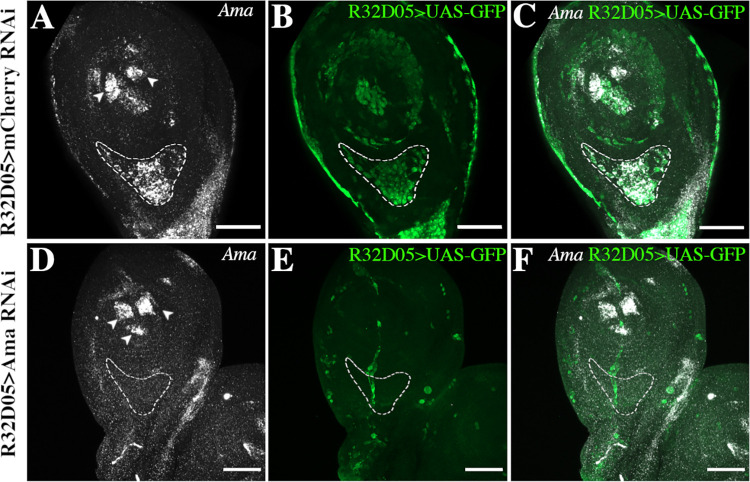
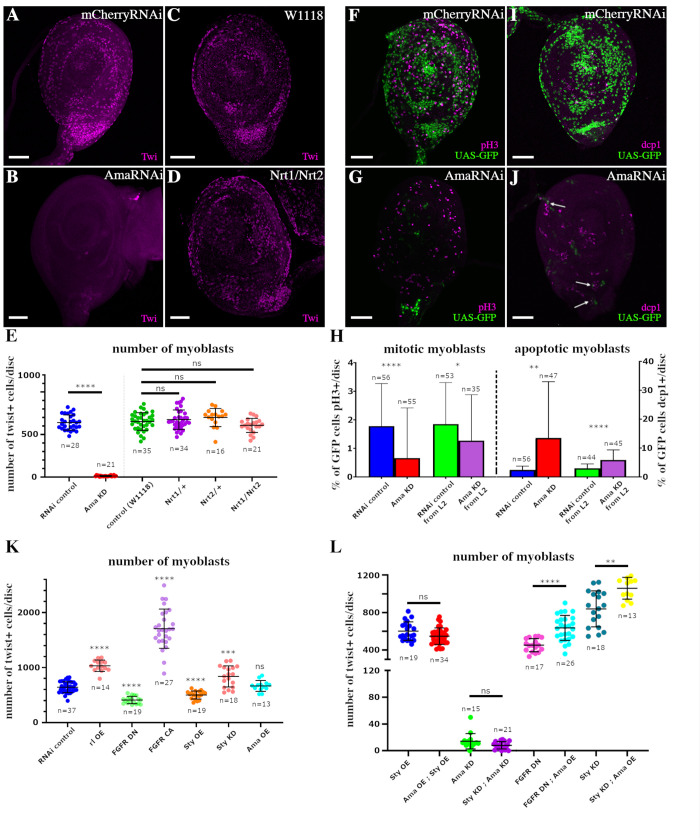
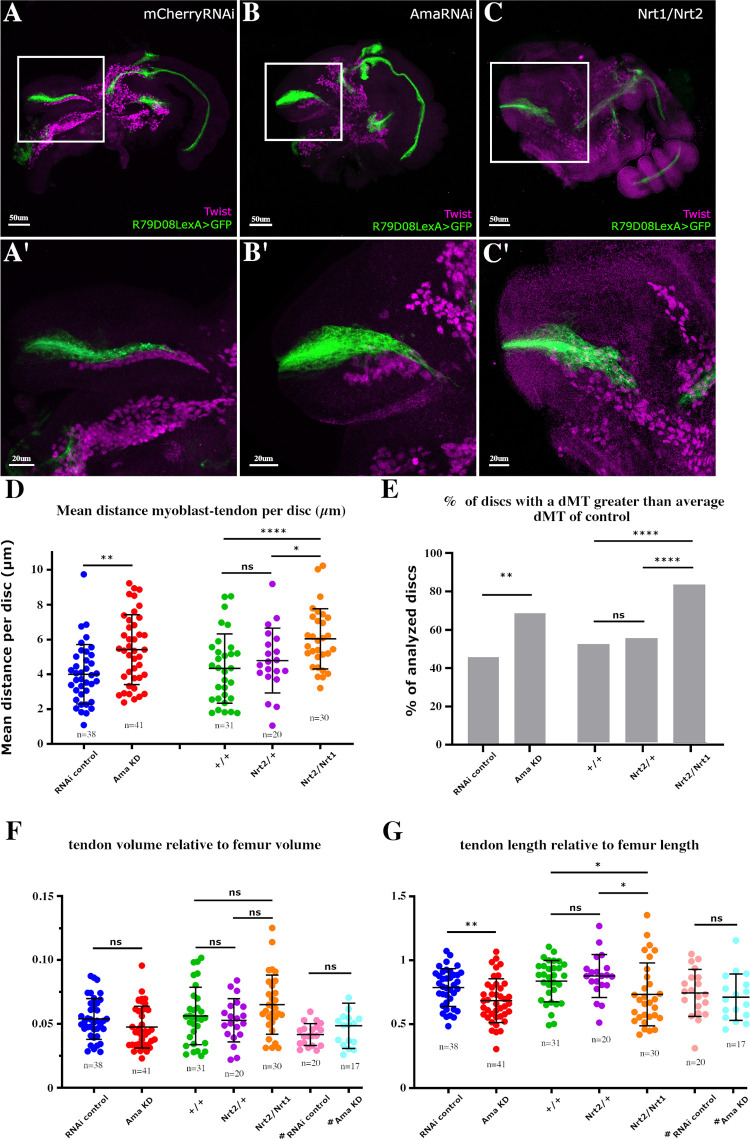
Formation of functional organs requires cell-cell communication between different cell lineages and failure in this communication can result in severe developmental defects. Hundreds of possible interacting pairs of proteins are known, but identifying the interacting partners that ensure a specific interaction between 2 given cell types remains challenging. Here, we use the Drosophila leg model and our cell type-specific transcriptomic data sets to uncover the molecular mediators of cell-cell communication between tendon and muscle precursors. Through the analysis of gene expression signatures of appendicular muscle and tendon precursor cells, we identify 2 candidates for early interactions between these 2 cell populations: Amalgam (Ama) encoding a secreted protein and Neurotactin (Nrt) known to encode a membrane-bound protein. Developmental expression and function analyses reveal that: (i) Ama is expressed in the leg myoblasts, whereas Nrt is expressed in adjacent tendon precursors; and (ii) in Ama and Nrt mutants, myoblast-tendon cell-cell association is lost, leading to tendon developmental defects. Furthermore, we demonstrate that Ama acts downstream of the FGFR pathway to maintain the myoblast population by promoting cell survival and proliferation in an Nrt-independent manner. Together, our data pinpoint Ama and Nrt as molecular actors ensuring early reciprocal communication between leg muscle and tendon precursors, a prerequisite for the coordinated development of the appendicular musculoskeletal system.
Copyright: © 2024 Moucaud et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
