

Role of Membrane Estrogen Receptor Alpha on the Positive Feedback of Estrogens on Kisspeptin and GnRH Neurons
- PMID: 39375032
- PMCID: PMC11520851
- DOI: 10.1523/ENEURO.0271-23.2024
Role of Membrane Estrogen Receptor Alpha on the Positive Feedback of Estrogens on Kisspeptin and GnRH Neurons
Abstract
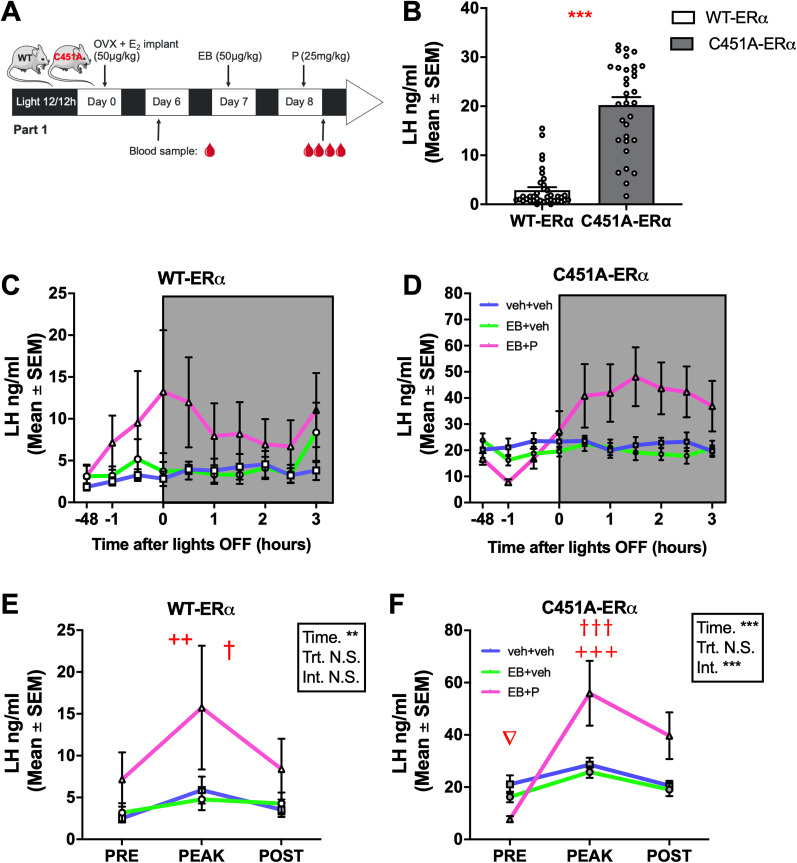
Estrogens act through nuclear and membrane-initiated signaling. Estrogen receptor alpha (ERα) is critical for reproduction, but the relative contribution of its nuclear and membrane signaling to the central regulation of reproduction is unclear. To address this question, two complementary approaches were used: estetrol (E4) a natural estrogen acting as an agonist of nuclear ERs, but as an antagonist of their membrane fraction, and the C451A-ERα mouse lacking mERα. E4 dose- dependently blocks ovulation in female rats, but the central mechanism underlying this effect is unknown. To determine whether E4 acts centrally to control ovulation, its effect was tested on the positive feedback of estradiol (E2) on neural circuits underlying luteinizing hormone (LH) secretion. In ovariectomized females chronically exposed to a low dose of E2, estradiol benzoate (EB) alone or combined with progesterone (P) induced an increase in the number of kisspeptin (Kp) and gonadotropin-releasing hormone (GnRH) neurons coexpressing Fos, a marker of neuronal activation. E4 blocked these effects of EB, but not when combined to P. These results indicate that E4 blocked the central induction of the positive feedback in the absence of P, suggesting an antagonistic effect of E4 on mERα in the brain as shown in peripheral tissues. In parallel, as opposed to wild-type females, C451A-ERα females did not show the activation of Kp and GnRH neurons in response to EB unless they are treated with P. Together these effects support a role for membrane-initiated estrogen signaling in the activation of the circuit mediating the LH surge.
Keywords: GnRH neurons; kisspeptin neurons; LH surge; estetrol; mERα; preoptic area.
Copyright © 2024 Faure et al.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources