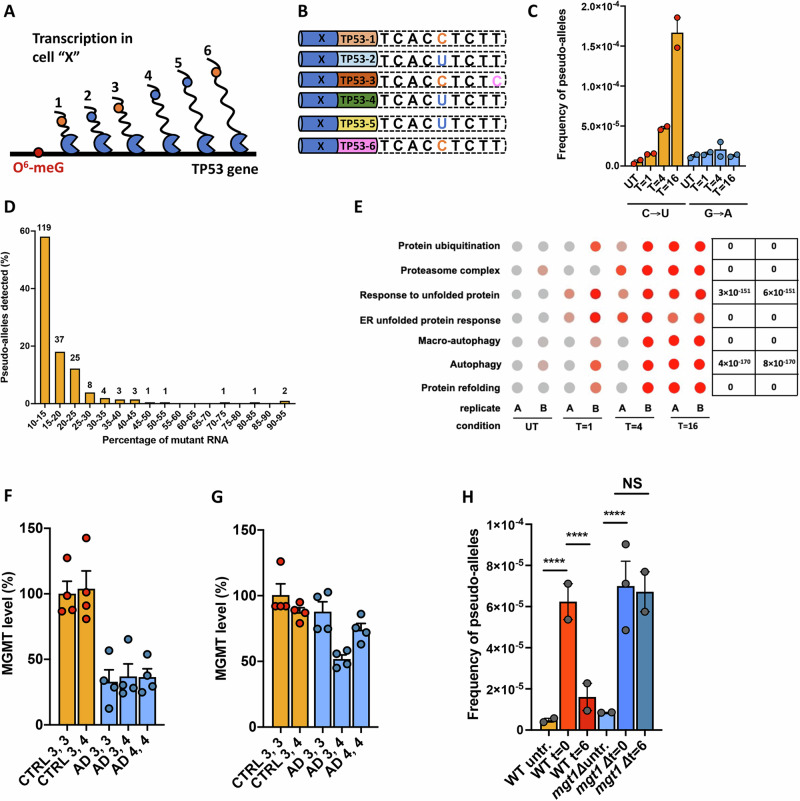
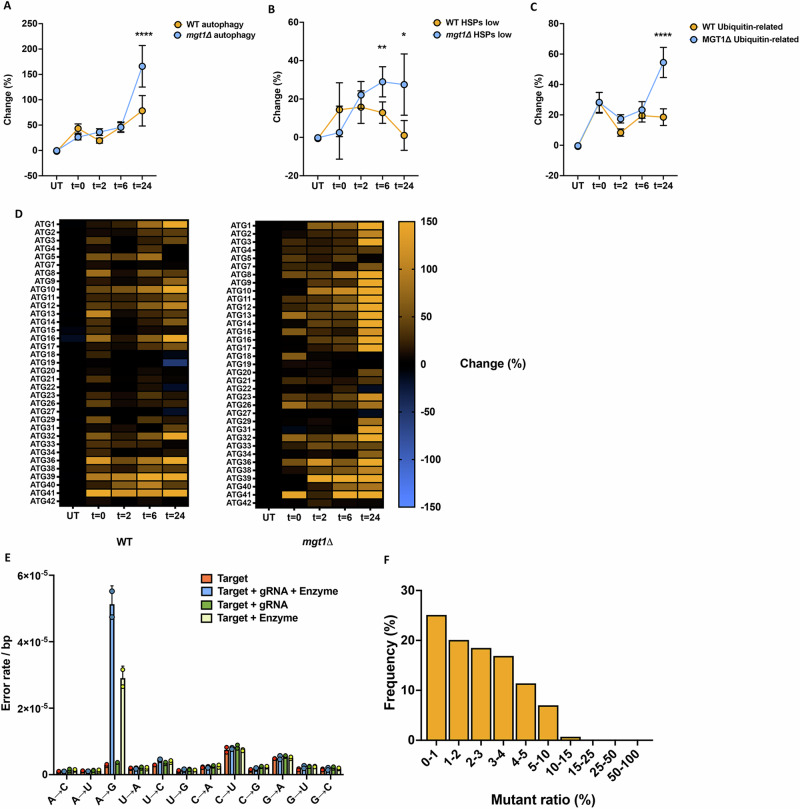
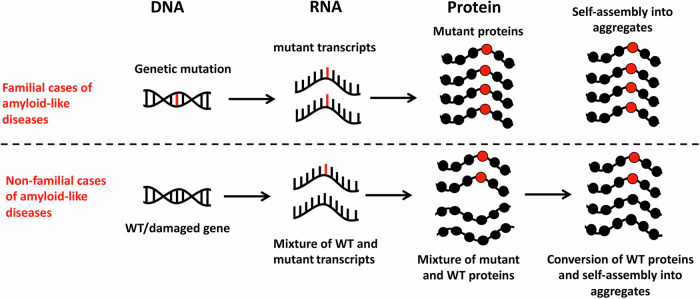
Transcript errors generate amyloid-like proteins in huwman cells
- PMID: 39375347
- PMCID: PMC11458900
- DOI: 10.1038/s41467-024-52886-2
Transcript errors generate amyloid-like proteins in huwman cells
Abstract
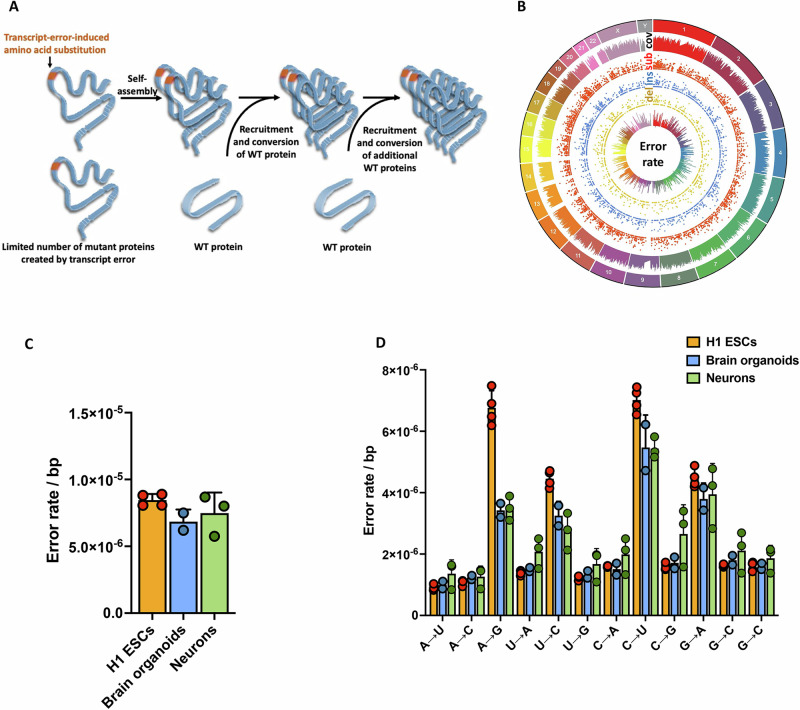
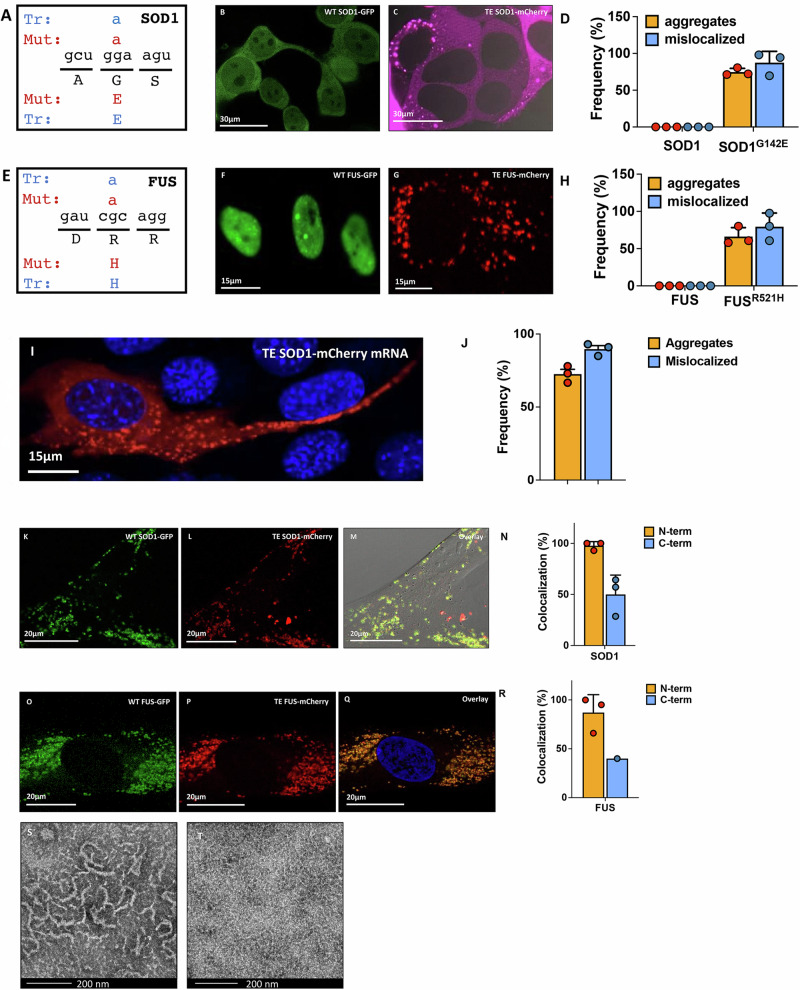
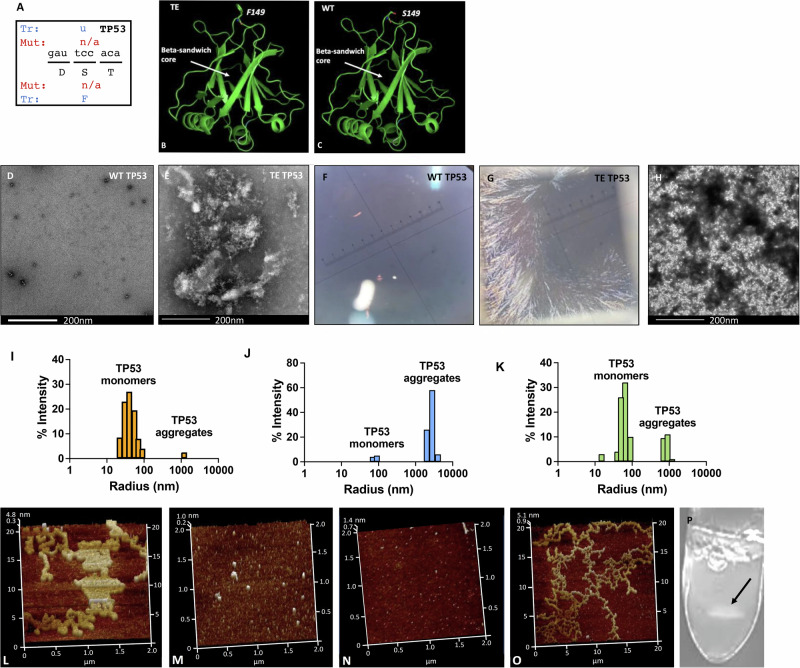
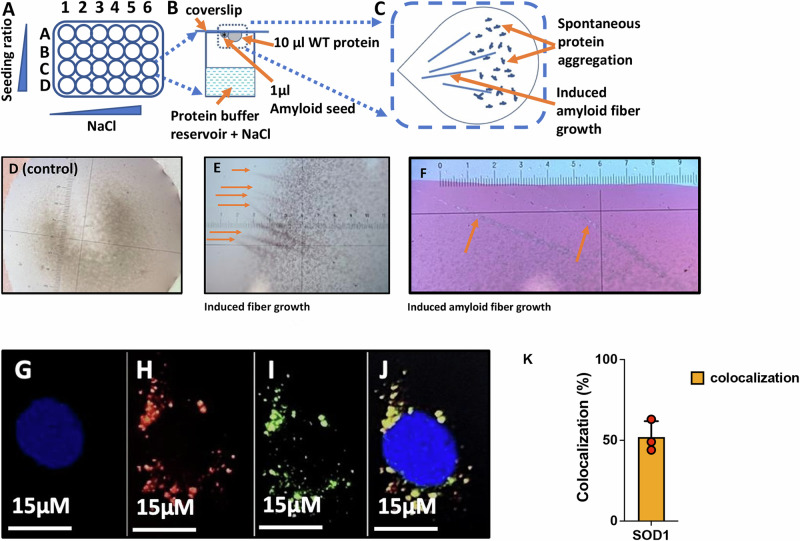
Aging is characterized by the accumulation of proteins that display amyloid-like behavior. However, the molecular mechanisms by which these proteins arise remain unclear. Here, we demonstrate that amyloid-like proteins are produced in a variety of human cell types, including stem cells, brain organoids and fully differentiated neurons by mistakes that occur in messenger RNA molecules. Some of these mistakes generate mutant proteins already known to cause disease, while others generate proteins that have not been observed before. Moreover, we show that these mistakes increase when cells are exposed to DNA damage, a major hallmark of human aging. When taken together, these experiments suggest a mechanistic link between the normal aging process and age-related diseases.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures