

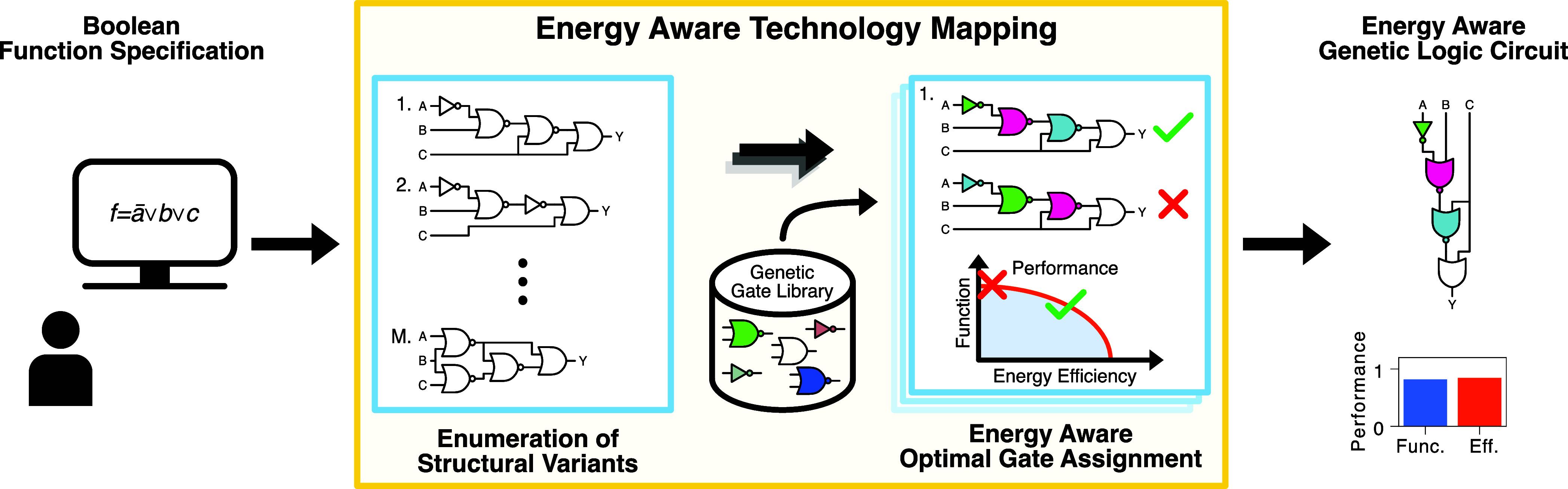
Energy Aware Technology Mapping of Genetic Logic Circuits
- PMID: 39378113
- PMCID: PMC11494706
- DOI: 10.1021/acssynbio.4c00395
Energy Aware Technology Mapping of Genetic Logic Circuits
Abstract
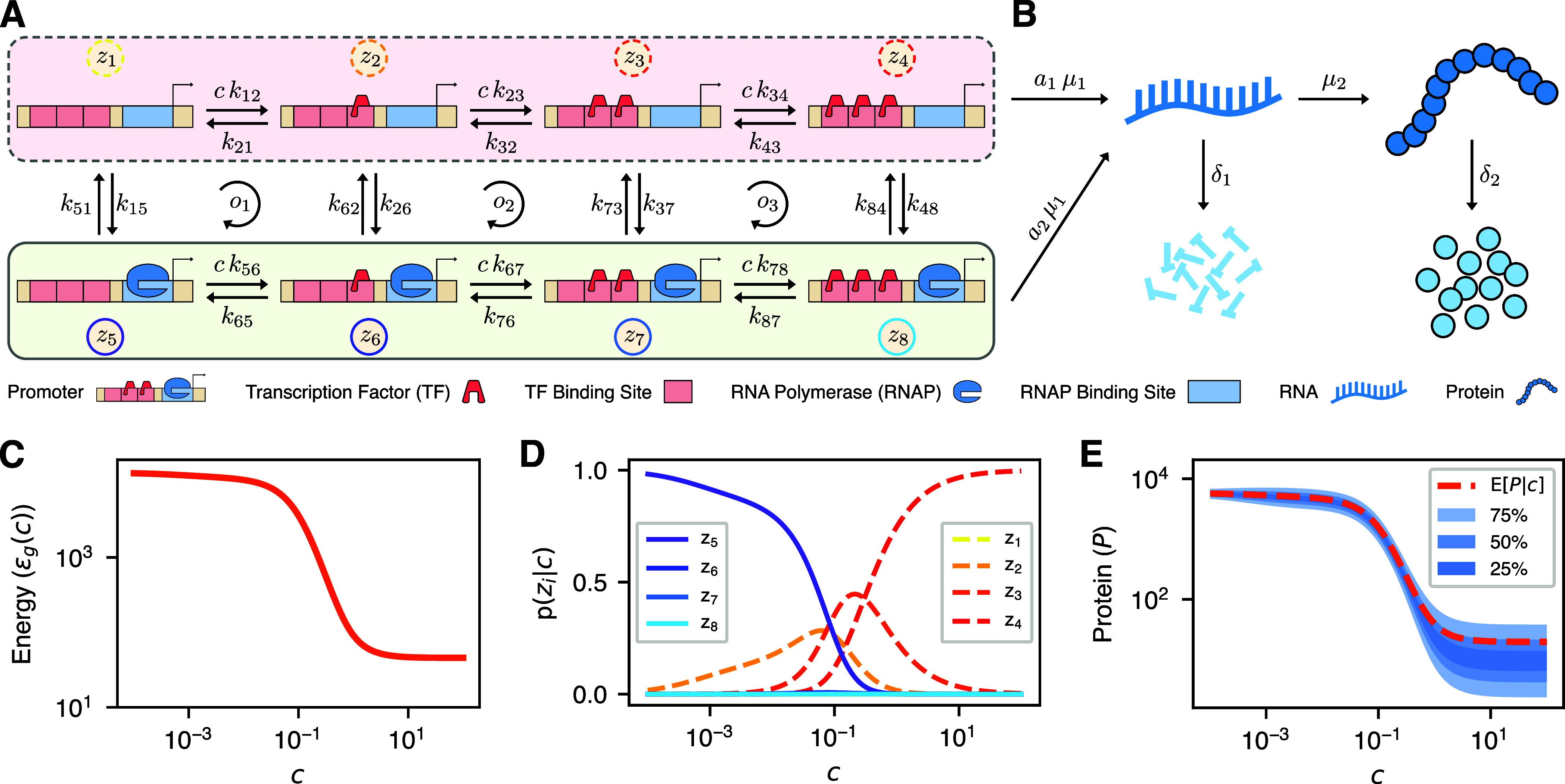
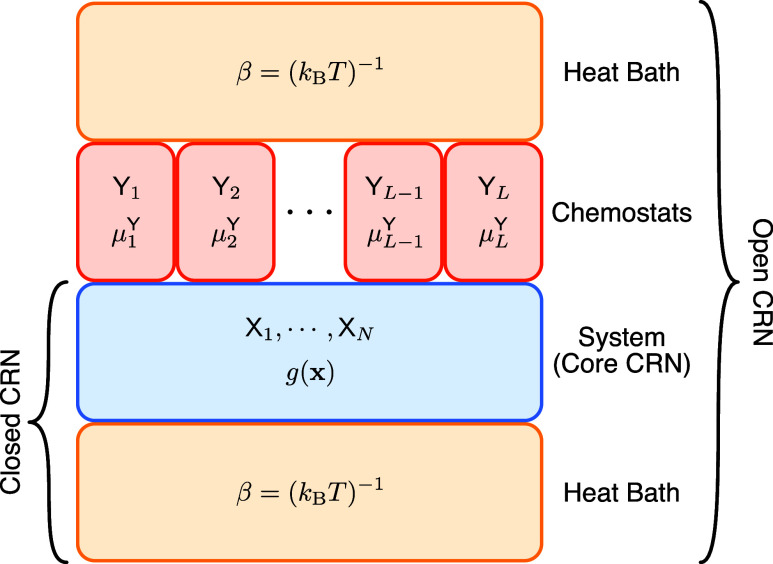
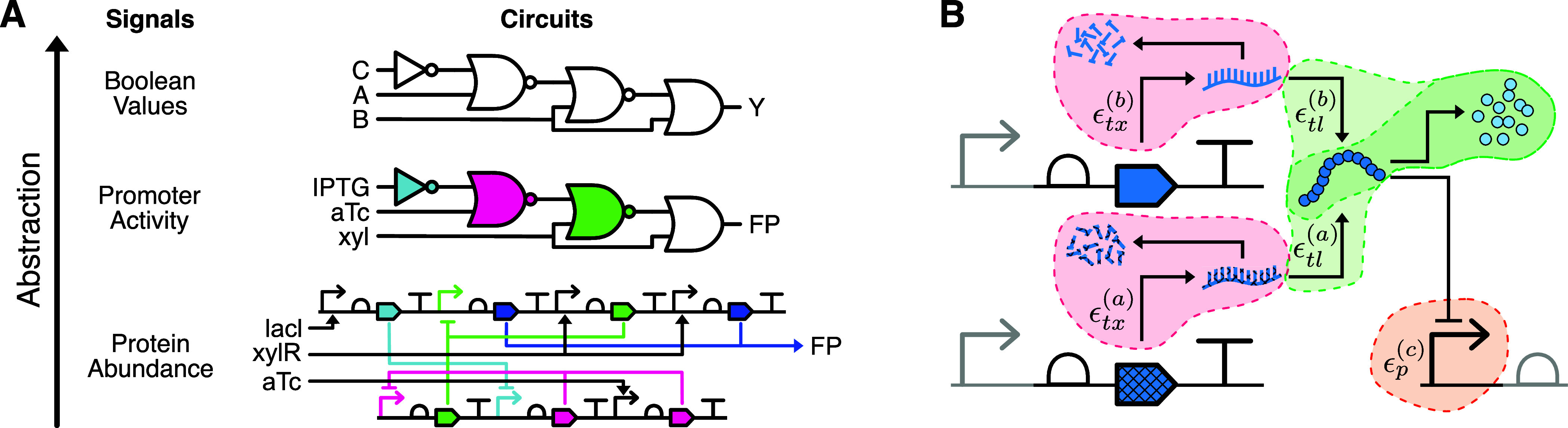
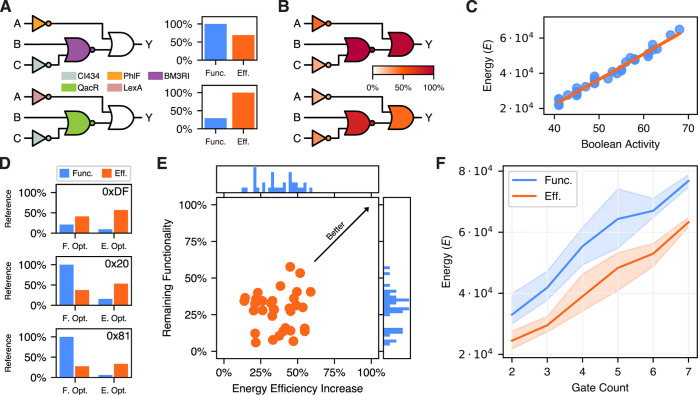
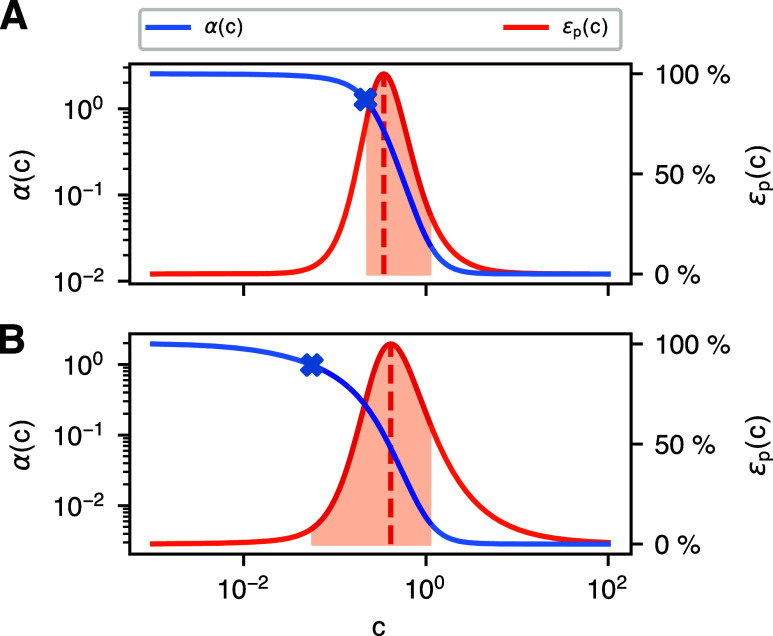
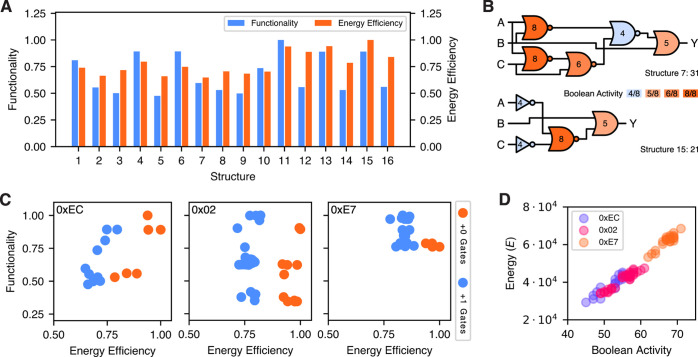
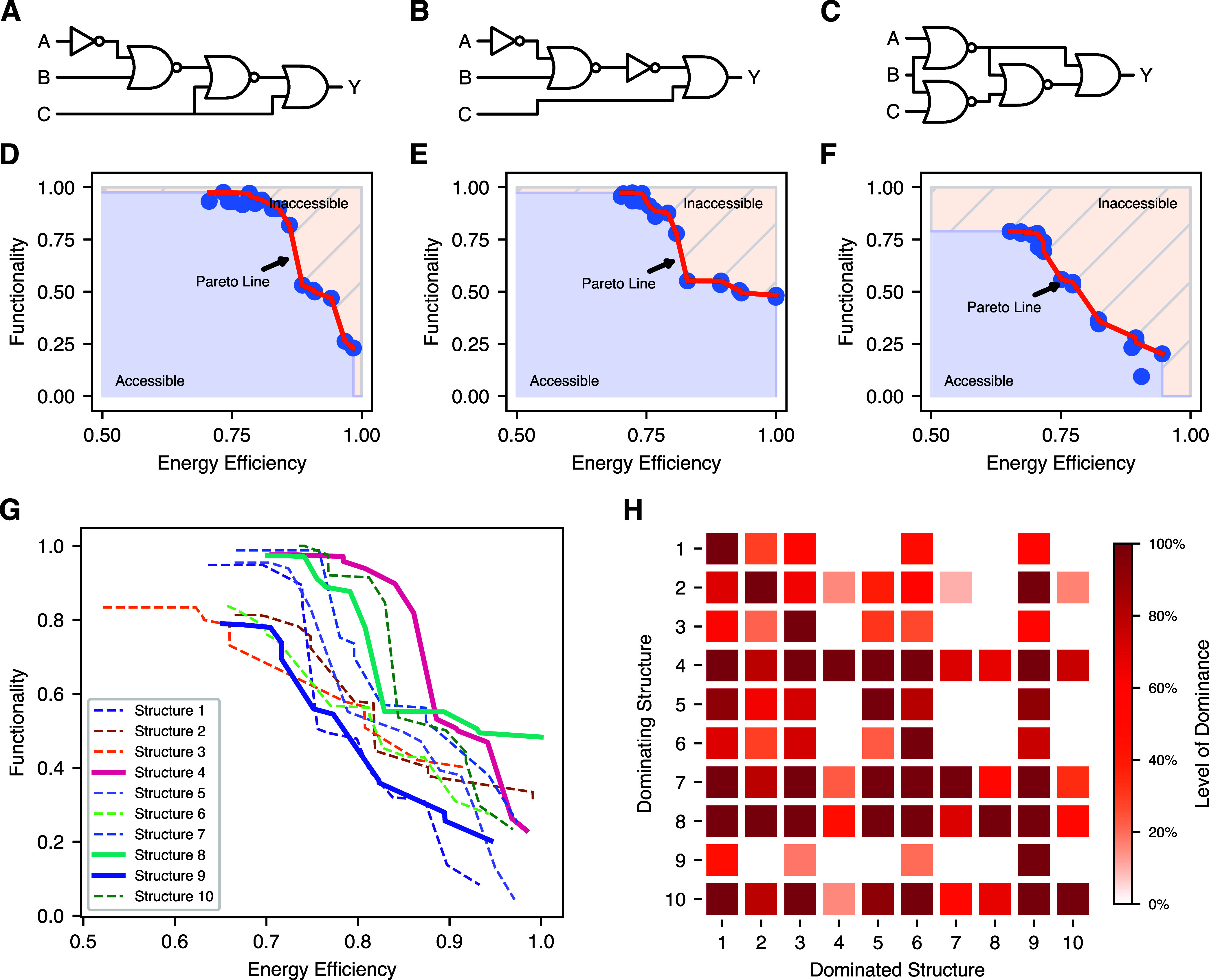
Energy and its dissipation are fundamental to all living systems, including cells. Insufficient abundance of energy carriers─as caused by the additional burden of artificial genetic circuits─shifts a cell's priority to survival, also impairing the functionality of the genetic circuit. Moreover, recent works have shown the importance of energy expenditure in information transmission. Despite living organisms being non-equilibrium systems, non-equilibrium models capable of accounting for energy dissipation and non-equilibrium response curves are not yet employed in genetic design automation (GDA) software. To this end, we introduce Energy Aware Technology Mapping, the automated design of genetic logic circuits with respect to energy efficiency and functionality. The basis for this is an energy aware non-equilibrium steady state model of gene expression, capturing characteristics like energy dissipation─which we link to the entropy production rate─and transcriptional bursting, relevant to eukaryotes as well as prokaryotes. Our evaluation shows that a genetic logic circuit's functional performance and energy efficiency are disjoint optimization goals. For our benchmark, energy efficiency improves by 37.2% on average when comparing to functionally optimized variants. We discover a linear increase in energy expenditure and overall protein expression with the circuit size, where Energy Aware Technology Mapping allows for designing genetic logic circuits with the energetic costs of circuits that are one to two gates smaller. Structural variants improve this further, while results show the Pareto dominance among structures of a single Boolean function. By incorporating energy demand into the design, Energy Aware Technology Mapping enables energy efficiency by design. This extends current GDA tools and complements approaches coping with burden in vivo.
Keywords: computer aided design; energy; entropy production rate; gene-expression; genetic design automation; metabolic burden; non-equilibrium; synthetic biology; technology mapping; thermodynamics.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Update of
-
Energy Aware Technology Mapping of Genetic Logic Circuits.bioRxiv [Preprint]. 2024 Sep 24:2024.06.27.601038. doi: 10.1101/2024.06.27.601038. bioRxiv. 2024. Update in: ACS Synth Biol. 2024 Oct 18;13(10):3295-3311. doi: 10.1021/acssynbio.4c00395. PMID: 39386604 Free PMC article. Updated. Preprint.
References
-
- Alberts B.; Johnson A.; Lewis J.; Raff M.; Roberts K.; Walter P.. Molecular Biology of the Cell, 6th ed.; Garland Science: New York, NY, 2017.
