

Multiomic single-cell sequencing defines tissue-specific responses in Stevens-Johnson syndrome and toxic epidermal necrolysis
- PMID: 39379371
- PMCID: PMC11461852
- DOI: 10.1038/s41467-024-52990-3
Multiomic single-cell sequencing defines tissue-specific responses in Stevens-Johnson syndrome and toxic epidermal necrolysis
Abstract
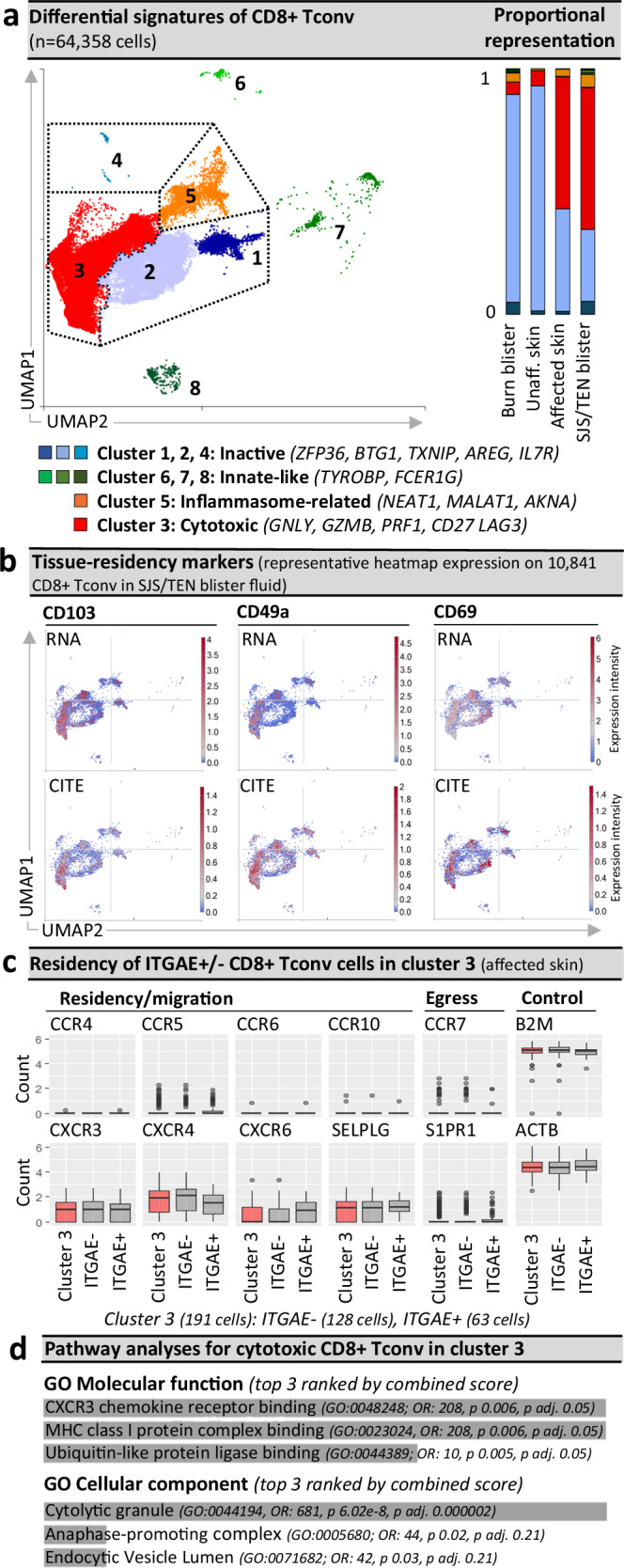
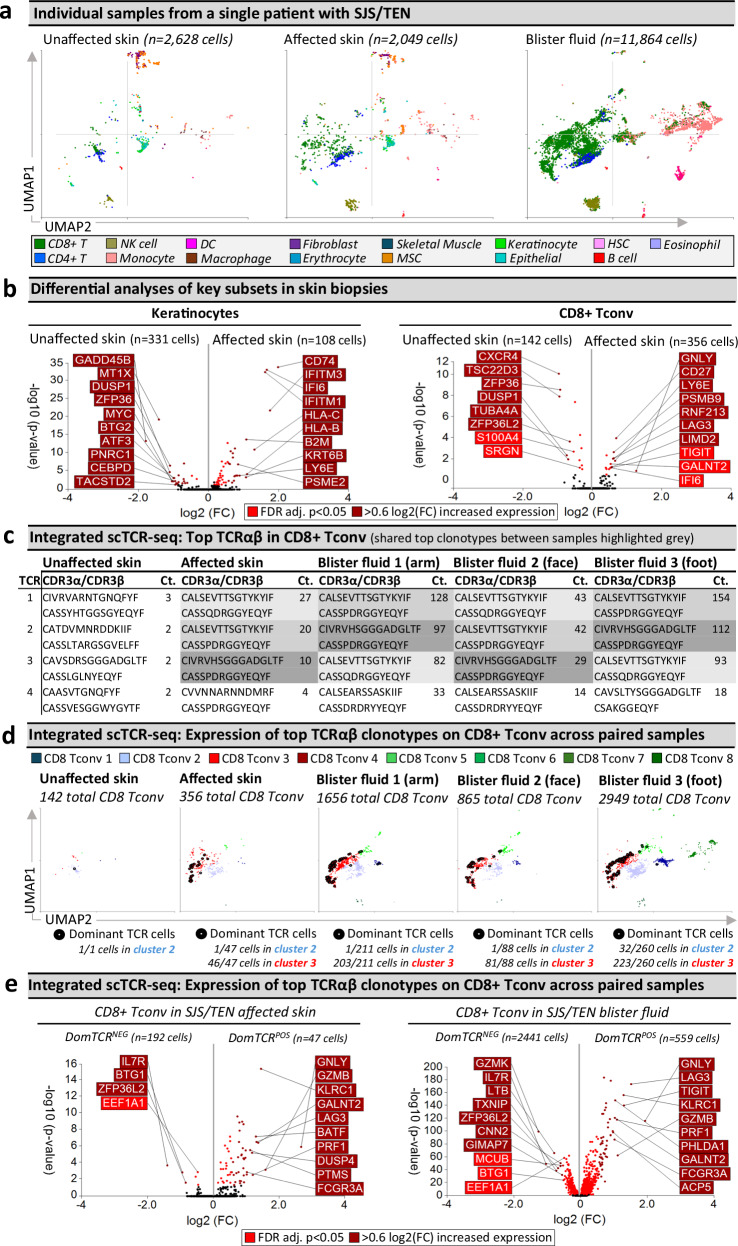
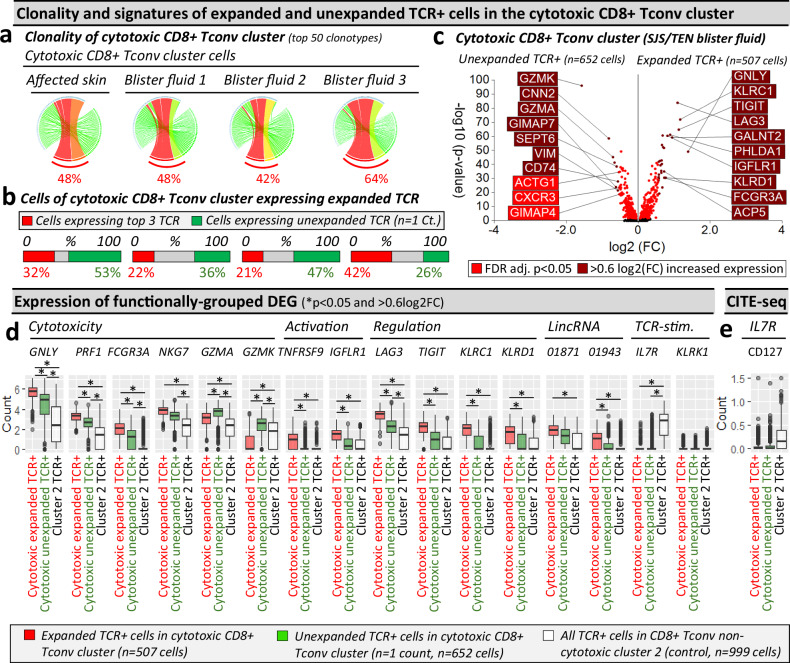
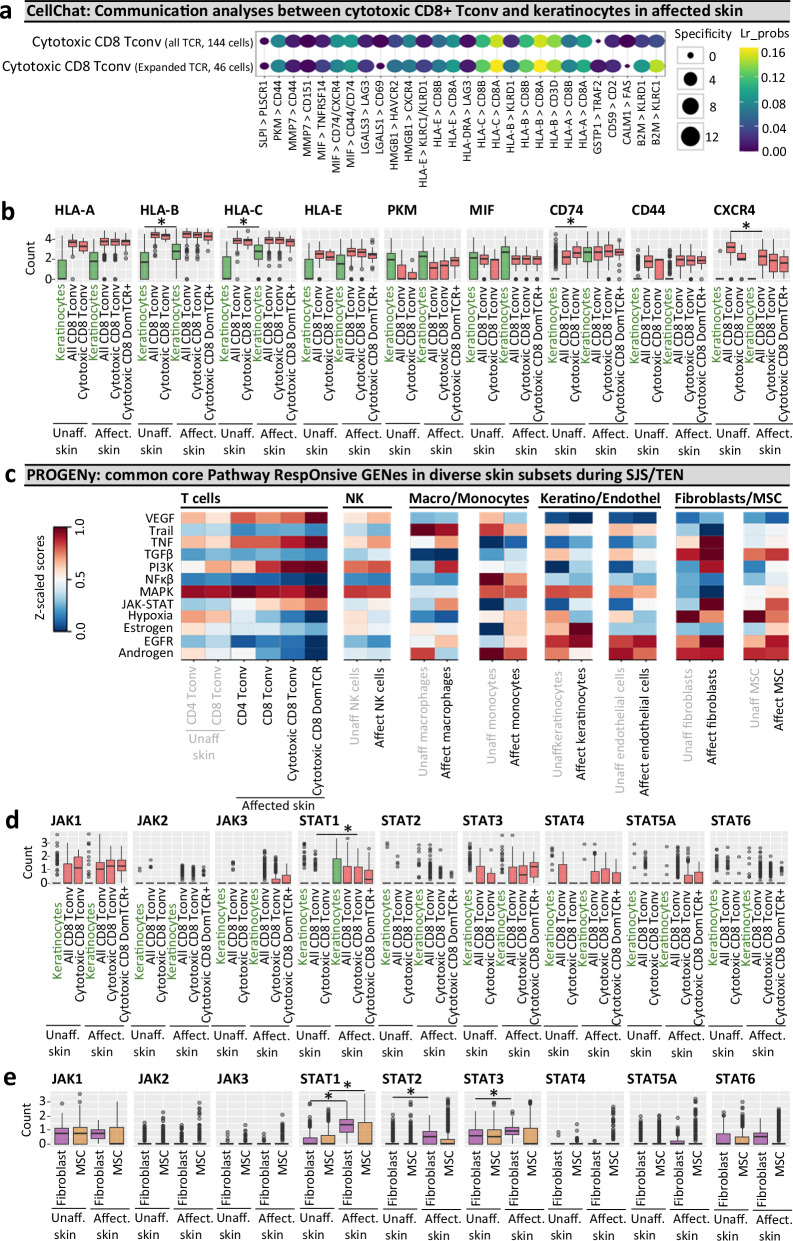
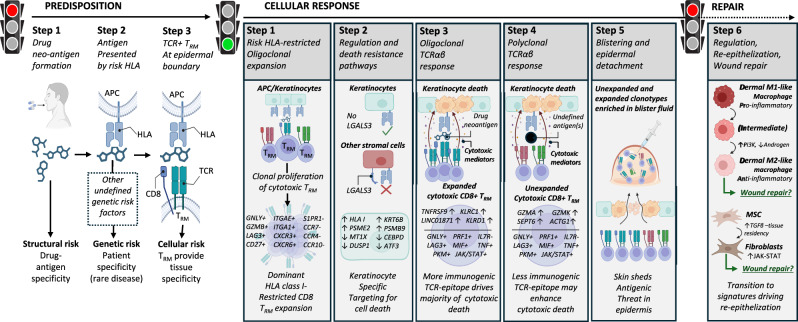
Stevens-Johnson syndrome and toxic epidermal necrolysis (SJS/TEN) is a rare but life-threatening cutaneous drug reaction mediated by human leukocyte antigen (HLA) class I-restricted CD8+ T cells. For unbiased assessment of cellular immunopathogenesis, here we perform single-cell (sc) transcriptome, surface proteome, and T cell receptor (TCR) sequencing on unaffected skin, affected skin, and blister fluid from 15 SJS/TEN patients. From 109,888 cells, we identify 15 scRNA-defined subsets. Keratinocytes express markers indicating HLA class I-restricted antigen presentation and appear to trigger the proliferation of and killing by cytotoxic CD8+ tissue-resident T cells that express granulysin, granzyme B, perforin, LAG3, CD27, and LINC01871, and signal through the PKM, MIF, TGFβ, and JAK-STAT pathways. In affected tissue, cytotoxic CD8+ T cells express private expanded and unexpanded TCRαβ that are absent or unexpanded in unaffected skin, and mixed populations of macrophages and fibroblasts express pro-inflammatory markers or those favoring repair. This data identifies putative cytotoxic TCRs and therapeutic targets.
© 2024. The Author(s).
Conflict of interest statement
E.J.P. receives royalties and consulting fees from UpToDate (where she is a Drug Allergy Section Editor) and has received consulting fees from Janssen, Vertex, Verve, Servier, Rapt and Esperion; she is co-director of IIID Pty Ltd, which holds a patent for HLA-B*57:01 testing for abacavir hypersensitivity, and she has a patent pending for detection of HLA-A*32:01 in connection with diagnosing drug reaction with eosinophilia and systemic symptoms (for these patents she does not receive any financial remuneration and neither are related to the submitted work). All other authors declare no competing interests.
Figures

Update of
-
Multiomic single-cell sequencing defines tissue-specific responses in Stevens-Johnson Syndrome and Toxic epidermal necrolysis.bioRxiv [Preprint]. 2024 Feb 18:2023.11.26.568771. doi: 10.1101/2023.11.26.568771. bioRxiv. 2024. Update in: Nat Commun. 2024 Oct 8;15(1):8722. doi: 10.1038/s41467-024-52990-3. PMID: 38405793 Free PMC article. Updated. Preprint.
References
-
- Krantz, M., Yoon, B., Stone, C., Yu, R. & Phillips, E. Stevens-Johnson syndrome and toxic epidermal necrolysis in the FDA adverse event reporting system (FAERS) from 1995-2020. J. Allergy Clin. Immunol.149, AB62 (2022).
-
- Posadas, S. J. et al. Delayed reactions to drugs show levels of perforin, granzyme B, and Fas-L to be related to disease severity. J. Allergy Clin. Immunol.109, 155–161 (2002). - PubMed
-
- Chung, W.-H. et al. Granulysin is a key mediator for disseminated keratinocyte death in Stevens-Johnson syndrome and toxic epidermal necrolysis. Nat. Med.14, 1343–1350 (2008). - PubMed
Publication types
MeSH terms
Substances
Associated data
- SRA/PRJNA1070820
- Actions
Grants and funding
- P50GM115305/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R13 AR082704/AR/NIAMS NIH HHS/United States
- R01AI152183/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R21AI139021/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 HG010863/HG/NHGRI NIH HHS/United States
- R01HG010863/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- D43 TW010559/TW/FIC NIH HHS/United States
- R01 AI152183/AI/NIAID NIH HHS/United States
- U01 AI154659/AI/NIAID NIH HHS/United States
- P30 AI110527/AI/NIAID NIH HHS/United States
- R21 AI139021/AI/NIAID NIH HHS/United States
- 2 D43 TW010559/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- GNT1123499/Department of Health | National Health and Medical Research Council (NHMRC)
- IK2 BX004585/BX/BLRD VA/United States
- K43 TW011178/TW/FIC NIH HHS/United States
- U01AI154659/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- GNT2028952/Department of Health | National Health and Medical Research Council (NHMRC)
- P50 GM115305/GM/NIGMS NIH HHS/United States
- K43 TW011178-01/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
