

Shade-induced ROS/NO reinforce COP1-mediated diffuse cell growth
- PMID: 39382994
- PMCID: PMC11494356
- DOI: 10.1073/pnas.2320187121
Shade-induced ROS/NO reinforce COP1-mediated diffuse cell growth
Abstract
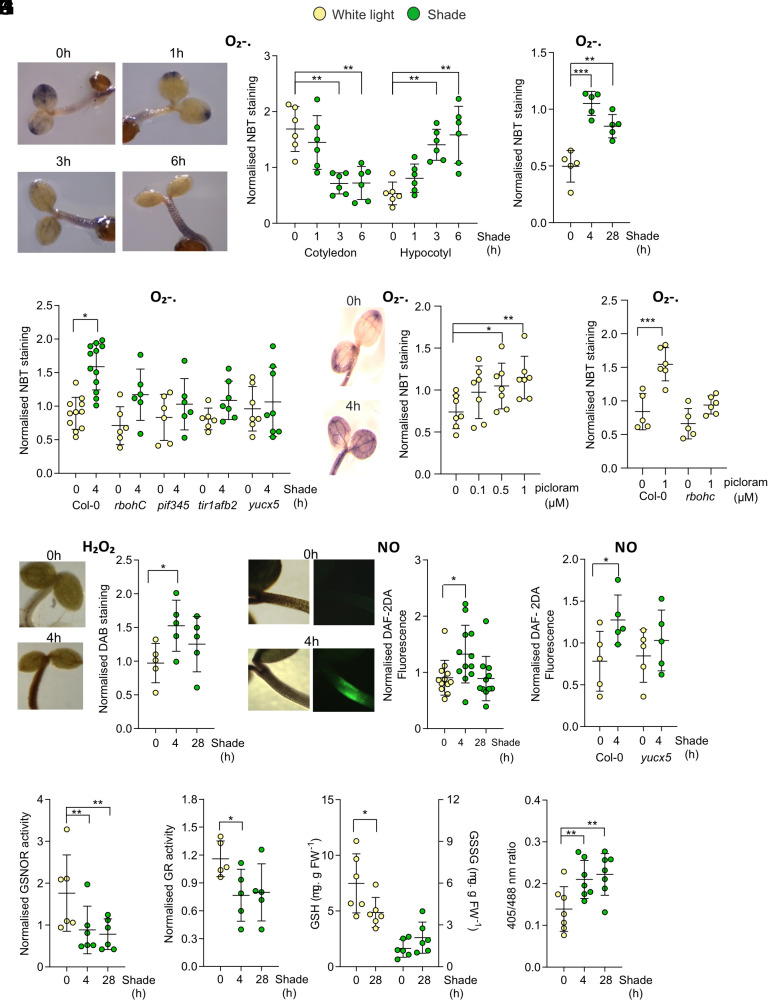
Canopy shade enhances the activity of PHYTOCHROME INTERACTING FACTORs (PIFs) to boost auxin synthesis in the cotyledons. Auxin, together with local PIFs and their positive regulator CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1), promotes hypocotyl growth to facilitate access to light. Whether shade alters the cellular redox status thereby affecting growth responses, remains unexplored. Here, we show that, under shade, high auxin levels increased reactive oxygen species and nitric oxide accumulation in the hypocotyl of Arabidopsis. This nitroxidative environment favored the promotion of hypocotyl growth by COP1 under shade. We demonstrate that COP1 is S-nitrosylated, particularly under shade. Impairing this redox regulation enhanced COP1 degradation by the proteasome and diminished the capacity of COP1 to interact with target proteins and to promote hypocotyl growth. Disabling this regulation also generated transversal asymmetries in hypocotyl growth, indicating poor coordination among different cells, which resulted in random hypocotyl bending and predictably low ability to compete with neighbors. These findings highlight the significance of redox signaling in the control of diffuse growth during shade avoidance.
Keywords: S-nitrosylation; auxin; hypocotyl growth; redox; shade avoidance.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures





References
-
- De Lucas M., et al. , A molecular framework for light and gibberellin control of cell elongation. Nature 451, 480–484 (2008). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
