

A potent pan-sarbecovirus neutralizing antibody resilient to epitope diversification
- PMID: 39383863
- PMCID: PMC11645210
- DOI: 10.1016/j.cell.2024.09.026
A potent pan-sarbecovirus neutralizing antibody resilient to epitope diversification
Abstract
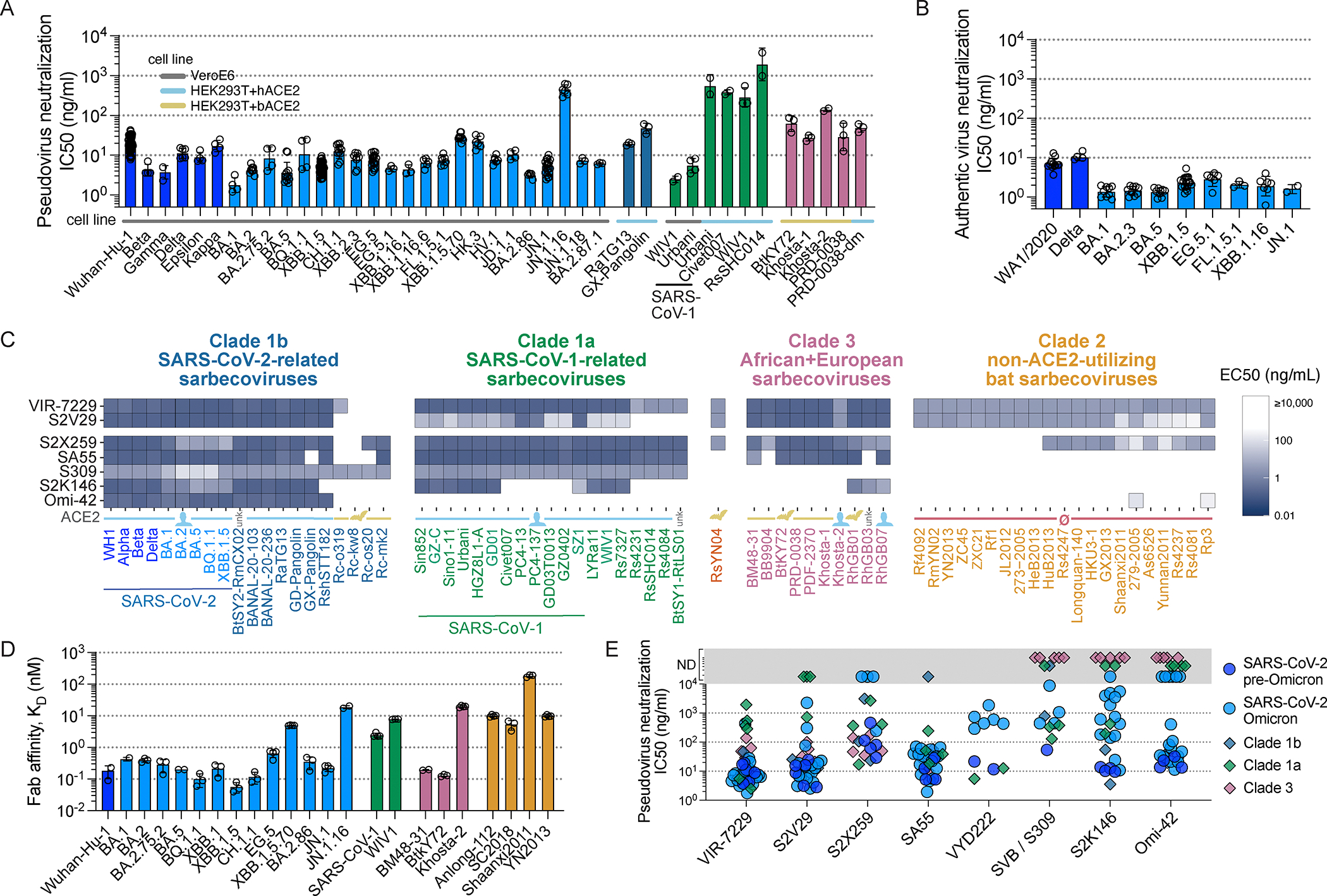
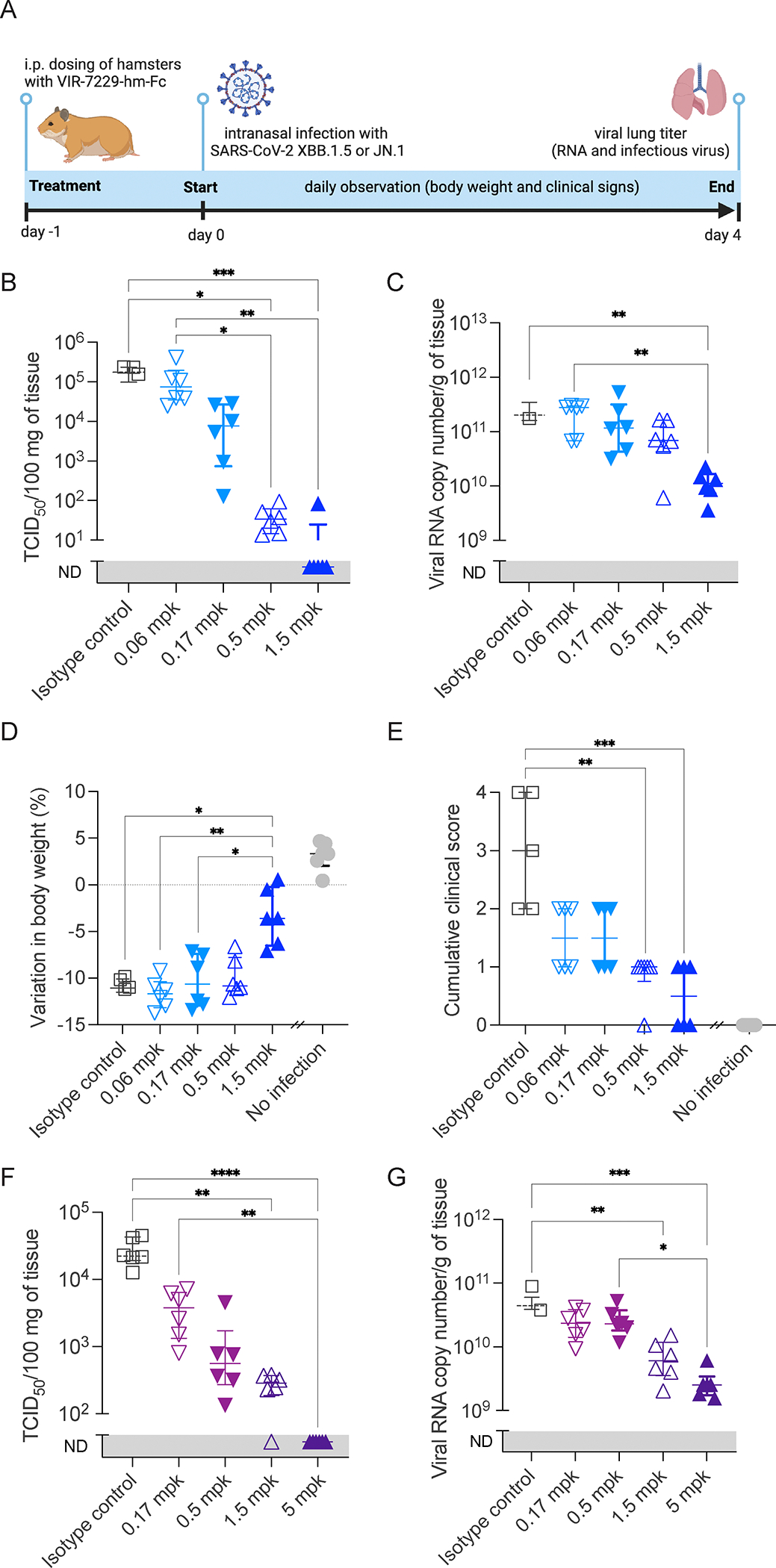
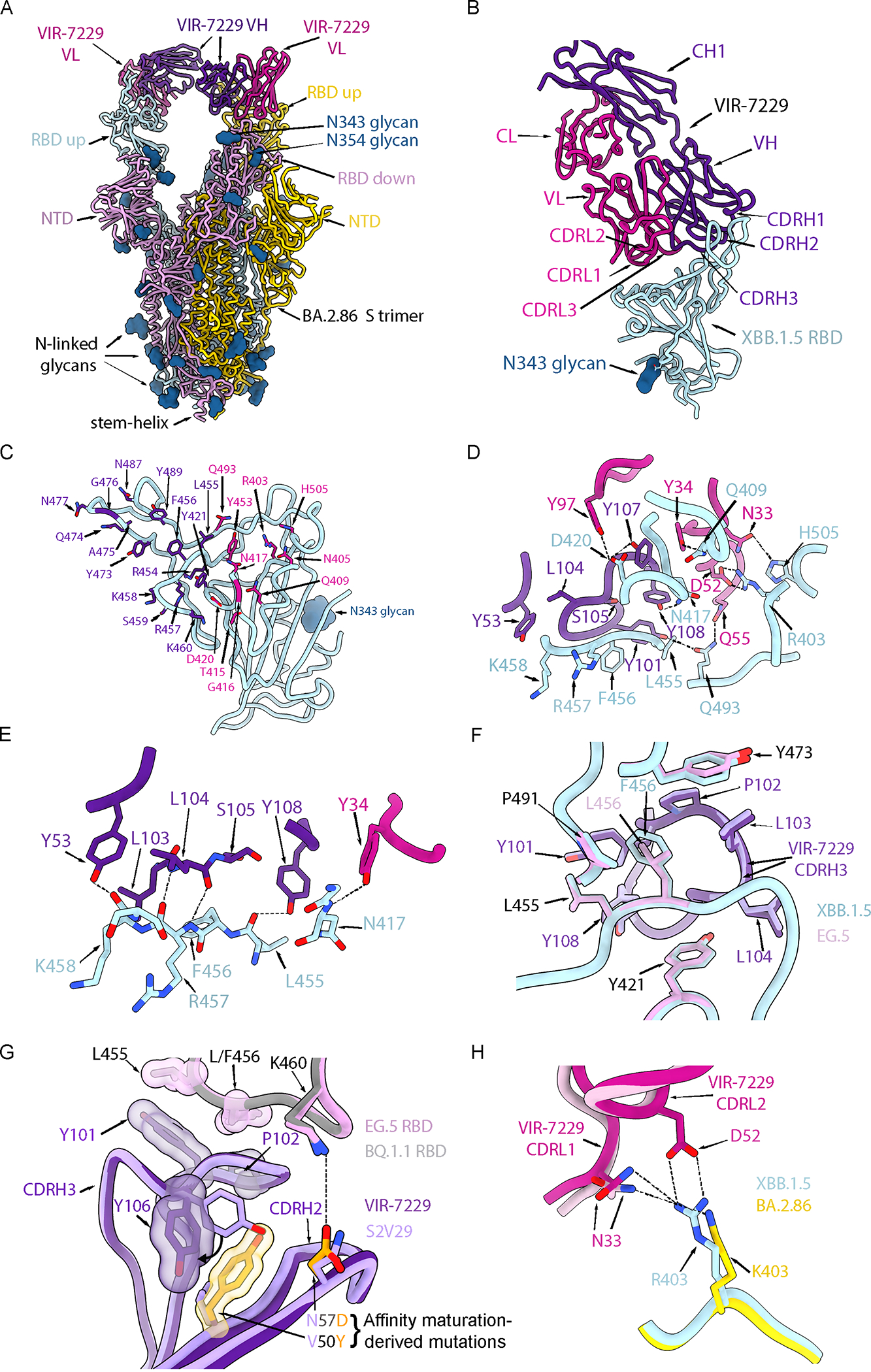
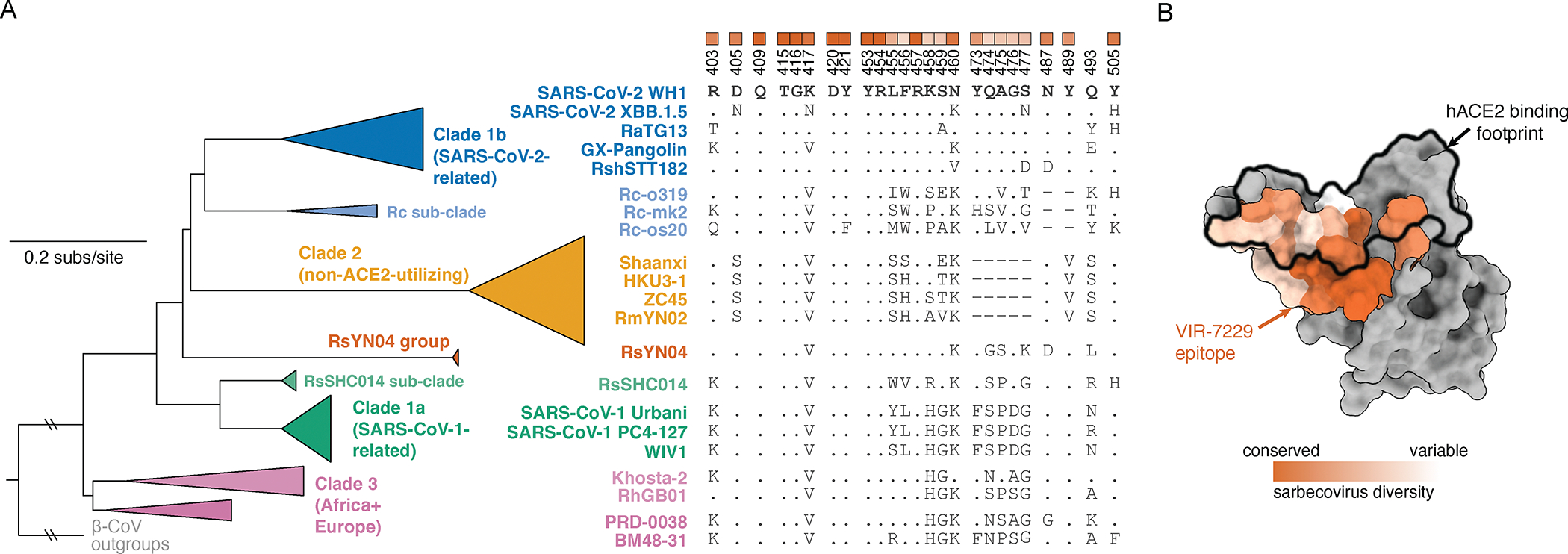
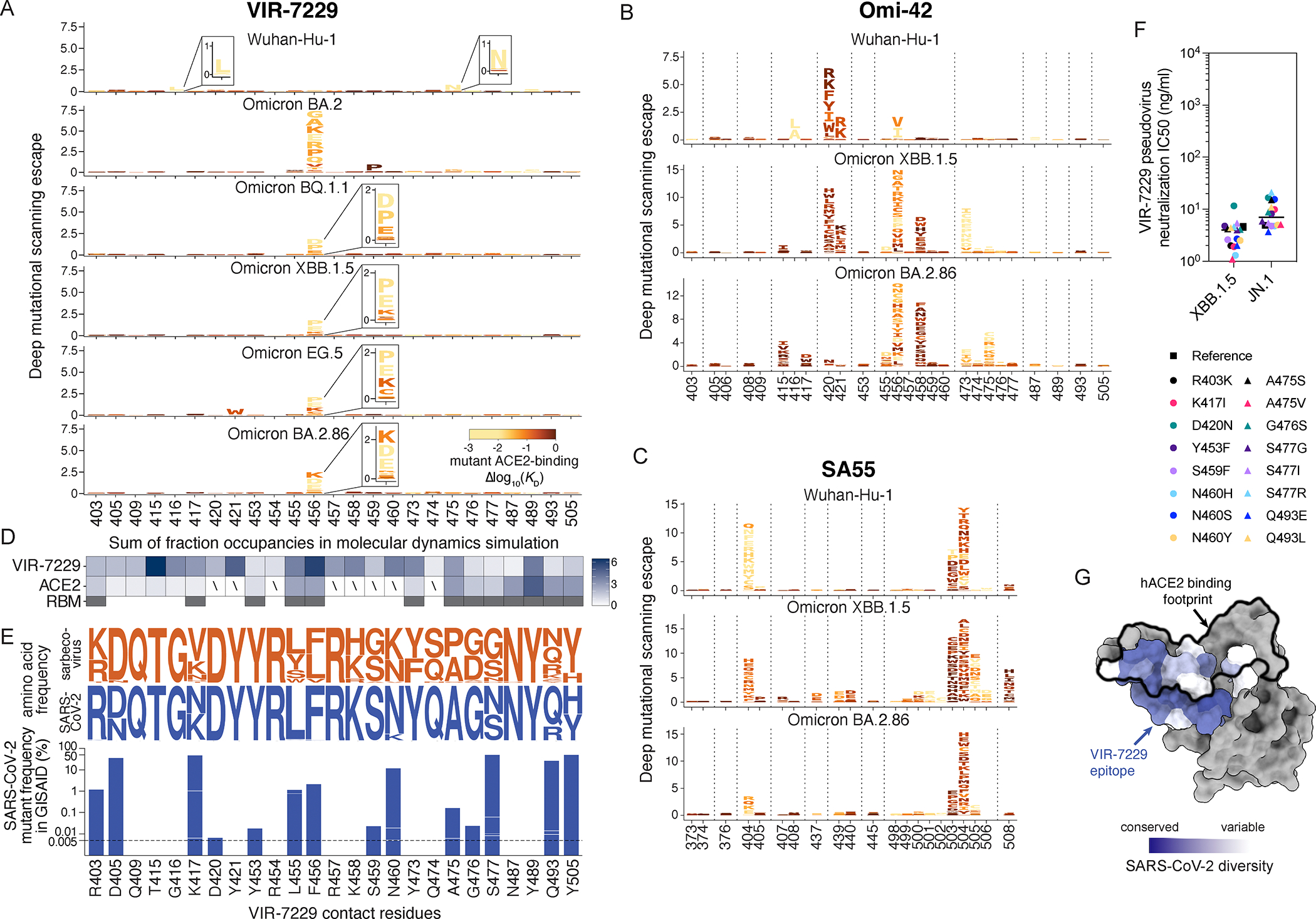
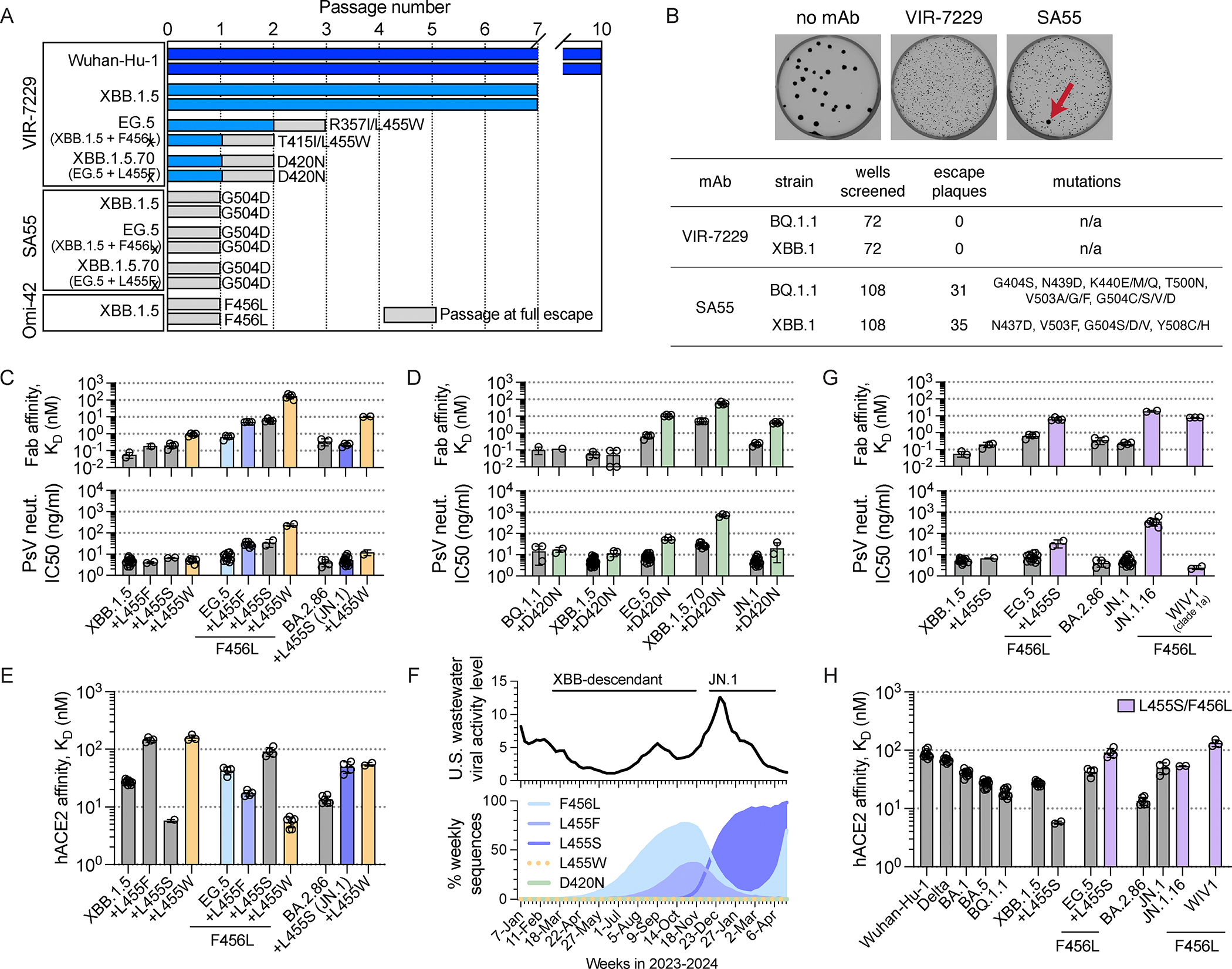
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) evolution has resulted in viral escape from clinically authorized monoclonal antibodies (mAbs), creating a need for mAbs that are resilient to epitope diversification. Broadly neutralizing coronavirus mAbs that are sufficiently potent for clinical development and retain activity despite viral evolution remain elusive. We identified a human mAb, designated VIR-7229, which targets the viral receptor-binding motif (RBM) with unprecedented cross-reactivity to all sarbecovirus clades, including non-ACE2-utilizing bat sarbecoviruses, while potently neutralizing SARS-CoV-2 variants since 2019, including the recent EG.5, BA.2.86, and JN.1. VIR-7229 tolerates extraordinary epitope variability, partly attributed to its high binding affinity, receptor molecular mimicry, and interactions with RBM backbone atoms. Consequently, VIR-7229 features a high barrier for selection of escape mutants, which are rare and associated with reduced viral fitness, underscoring its potential to be resilient to future viral evolution. VIR-7229 is a strong candidate to become a next-generation medicine.
Keywords: SARS-CoV-2; broadly neutralizing antibody; monoclonal antibody; sarbecovirus; viral antibody escape.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L.E.R., A.D.M., D.P., D.B., T.R., J.M.E., K.H., H.V.D., M.G., G.C., C.S., F.Z., E.D., A.R., J.N., Q.T., Y.D., B.T., R.A., S.S., B.G., M.A.S., G. Schnell, J.L.M., F.A.L., N.C., E.C., L.A.P., F.B., J.d.I., M.S.P., A.L., A.T., G. Snell, and D.C. are current or previous employees of Vir Biotechnology and may hold shares in Vir Biotechnology. L.E.R., A.D.M., D.P., E.C., F.B., M.S.P., G. Snell, and D.C. are currently listed as inventors on multiple patent applications that disclose the subject matter described in this paper. J.W.C. is an employee and shareholder of ProtaBody. J.W.C. and ProtaBody have received funding from Vir Biotechnology related to the work described in this paper. I.Z., Z.L., S.P.J.W., G.D.d.M., L.K., H.B., and T.N.S. have received funding through sponsored research awards to their respective institutions from Vir Biotechnology related to the work described in this paper. I.Z. is a current employee of Bristol Myers Squibb. L.A.P. is a former employee and shareholder of Regeneron Pharmaceuticals and is a member of the Scientific Advisory Board AI-driven structure-enabled antiviral platform (ASAP). Regeneron provided no funding for this work. L.A.P. is a current employee of Third Rock Ventures. D.V. is named as inventor on patents for coronavirus vaccines filed by the University of Washington. The lab of T.N.S. has received sponsored research agreements unrelated to the present work from Aerium Therapeutics, Inc. and Invivyd, Inc.
Figures
Comment in
-
Antibody vanquishes SARS-CoV-2 variants.Nat Rev Drug Discov. 2024 Dec;23(12):895. doi: 10.1038/d41573-024-00173-6. Nat Rev Drug Discov. 2024. PMID: 39472733 No abstract available.
References
-
- Suribhatla R, Starkey T, Ionescu MC, Pagliuca A, Richter A, and Lee LYW (2023). Systematic review and meta-analysis of the clinical effectiveness of tixagevimab/cilgavimab for prophylaxis of COVID-19 in immunocompromised patients. Br. J. Haematol. 201, 813–823. - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
