

Redox control of the deubiquitinating enzyme Ubp2 regulates translation during stress
- PMID: 39384040
- PMCID: PMC11570842
- DOI: 10.1016/j.jbc.2024.107870
Redox control of the deubiquitinating enzyme Ubp2 regulates translation during stress
Abstract
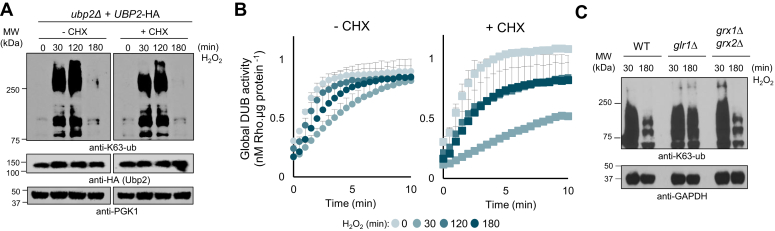
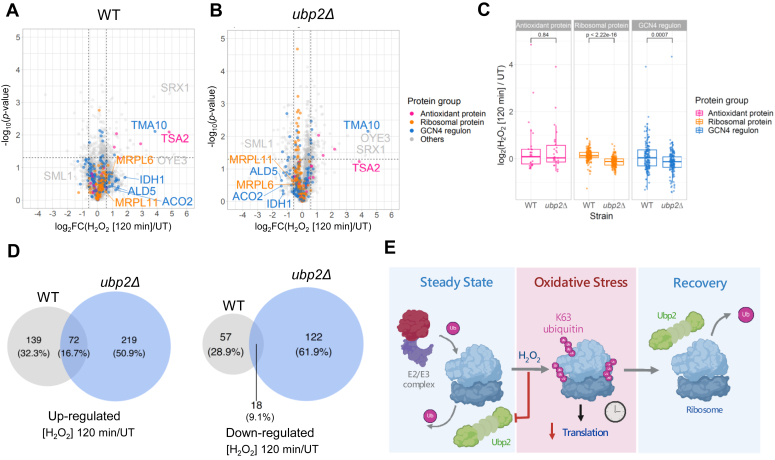
Protein ubiquitination is essential to govern cells' ability to cope with harmful environments by regulating many aspects of protein dynamics from synthesis to degradation. As important as the ubiquitination process, the reversal of ubiquitin chains mediated by deubiquitinating enzymes (DUBs) is critical for proper recovery from stress and re-establishment of proteostasis. Although it is known that ribosomes are decorated with K63-linked polyubiquitin chains that control protein synthesis under stress, the mechanisms by which these ubiquitin chains are reversed and regulate proteostasis during stress recovery remain elusive. Here, we showed in budding yeast that the DUB Ubp2 is redox-regulated during oxidative stress in a reversible manner, which determines the levels of K63-linked polyubiquitin chains present on ribosomes. We also demonstrate that Ubp2 can cleave single ubiquitin moieties out of chains and its activity is modulated by a series of repeated domains and the formation of disulfide bonds. By combining cellular, biochemical, and proteomics analyses, we showed that Ubp2 is crucial for restoring translation after stress cessation, indicating an important role in determining the cellular response to oxidative stress. Our work demonstrates a novel role for Ubp2, revealing that a range of signaling pathways can be controlled by redox regulation of DUB activity in eukaryotes, which in turn will define cellular states of health and diseases.
Keywords: deubiquitylation (deubiquitination); oxidative stress; redox regulation; translation control; yeast.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
