

Temporally distinct 3D multi-omic dynamics in the developing human brain
- PMID: 39385032
- PMCID: PMC11560841
- DOI: 10.1038/s41586-024-08030-7
Temporally distinct 3D multi-omic dynamics in the developing human brain
Abstract
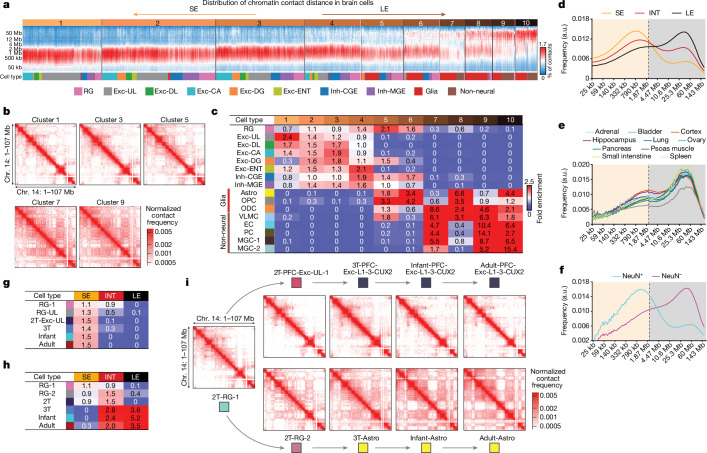
The human hippocampus and prefrontal cortex play critical roles in learning and cognition1,2, yet the dynamic molecular characteristics of their development remain enigmatic. Here we investigated the epigenomic and three-dimensional chromatin conformational reorganization during the development of the hippocampus and prefrontal cortex, using more than 53,000 joint single-nucleus profiles of chromatin conformation and DNA methylation generated by single-nucleus methyl-3C sequencing (snm3C-seq3)3. The remodelling of DNA methylation is temporally separated from chromatin conformation dynamics. Using single-cell profiling and multimodal single-molecule imaging approaches, we have found that short-range chromatin interactions are enriched in neurons, whereas long-range interactions are enriched in glial cells and non-brain tissues. We reconstructed the regulatory programs of cell-type development and differentiation, finding putatively causal common variants for schizophrenia strongly overlapping with chromatin loop-connected, cell-type-specific regulatory regions. Our data provide multimodal resources for studying gene regulatory dynamics in brain development and demonstrate that single-cell three-dimensional multi-omics is a powerful approach for dissecting neuropsychiatric risk loci.
© 2024. The Author(s).
Conflict of interest statement
Figures














References
MeSH terms
Substances
Grants and funding
- U01 MH116438/MH/NIMH NIH HHS/United States
- R01 NS123263/NS/NINDS NIH HHS/United States
- R01 HG010634/HG/NHGRI NIH HHS/United States
- DP2 NS122550/NS/NINDS NIH HHS/United States
- U01 MH121282/MH/NIMH NIH HHS/United States
- DP5 OD031878/OD/NIH HHS/United States
- U24 HG002371/HG/NHGRI NIH HHS/United States
- P01 NS083513/NS/NINDS NIH HHS/United States
- U01 MH130995/MH/NIMH NIH HHS/United States
- R01 MH125252/MH/NIMH NIH HHS/United States
- RF1 MH132662/MH/NIMH NIH HHS/United States
- R01 DA059602/DA/NIDA NIH HHS/United States
- R01 MH115676/MH/NIMH NIH HHS/United States
- R01 MH125516/MH/NIMH NIH HHS/United States
- UM1 MH130994/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
